تشابه المكونات 2: الحمض النووى ج1
- Illidan Stormrage

- Mar 7
- 57 min read
Updated: Mar 8
The unity of life is no less remarkable than its diversity. Most forms of life are similar in many respects. The universal biologic similarities are particularly striking in the biochemical dimension. From viruses to man, heredity is coded in just two, chemically related substances: DNA and RNA...striking uniformities prevail in the cellular metabolism of the most diverse living beings. Adenosine triphosphate, biotin, riboflavin, hemes, pyridoxin, vitamins K and B12, and folic acid implement metabolic processes everywhere.
What do these biochemical or biologic universals mean? They suggest that life arose from inanimate matter only once and that all organisms, no matter now diverse, in other respects, conserve the basic features of the primordial life. (It is also possible that there were several, or even many, origins of life; if so, the progeny of only one of them has survived and inherited the earth.) But what if there was no evolution and every one of the millions of species were created by separate fiat? However offensive the notion may be to religious feeling and to reason, the anti-evolutionists must again accuse the Creator of cheating. They must insist that He deliberately arranged things exactly as if his method of creation was evolution, intentionally to mislead sincere seekers of truth.
Theodosius Dobzhansky, "Nothing in biology makes sense except in the light of evolution." The American Biology Teacher, March 1973
فى مقالته الرائعة لا شئ فى البيولوجيا يمكن فهمه الا فى ضوء التطور يقدم لنا ثيودوسيوس دوبزانسكى ملخصا لأحد أهم أساليب الاستدلال اللاهوتية التطورية الملتوية – يعدد لنا دوبزانسكى تشابهات الكائنات الحية فى مكوناتها ابتداءا من الحمض النووى و مرورا بمركبات بيوكيميائية أخرى كثيرة لينهى العرض بالاستنتاج الحاسم: كل هذا يرجح أن الحياة نشأت من المادة الغير حية بهذه الصفات ثم تشعبت حاملة هذه السمات من ذلك السلف المشترك الذى نجا و استمر.
الان ركز معى فى عملية النصب العظمى التى حدثت لأنها غالبا ما سيفعله معك أى تطورى حتى لو لم يكن يحدثك عن أصل الحياة. لقد تم تجاوز استحالة نشأة الحياة من المادة الغير جية و التفاعلات الكيميائية و تجاوز عدم وجود أى الية مفصلة بخطوات وظيفية مثبتة لانتاج كل التعقيد الجينى/البيوكيميائى/التشريحى المعاصر و اعتبار حدوث هذه الأشياء تحصيل حاصل و القفز مباشرة الى حقيقة حدوث التطور. لهذا يكرر كثير من التطورييين جملة "التطور حقيقة حتى لو لم نفهم كيف يعمل"لأنه ببساطة لا يعمل أصلا و هم يهربون من هذا باعتبار التشابهات علامة حدوثه. و الدليل؟ حجة لاهوتية بامتياز اذ يكمل دوبزانسكى موضحا أن البديل عن هذا التصور هو أن الخالق يخدعنا ليوحى لنا بالتطور!
صديقنا التطورى الذى ان حدثته عن التصميم سيؤكد لك أن هذا ليس علما لأننا غير قادرين على ادخال الخالق/المصمم الى المعمل أو وضعه فى أنبوب اختبار فجأة أدخل الخالق المعمل و وضع التصميم فى أنبوب الاختبار و أجرى litmus test أكد له أن المصممين لا يستخدمون نفس المكونات أبدا! و بهذا يؤكد لنا من جديد صحة ما نقوله عن نظرية التطور دائما Evolutionary Theology not Biology انها لاهوت لتفسير علم الأحياء و ليست نظرية علمية. طبعا رجل القش الشهير كان حاضرا بقوة فى عرض الحجة فالمصمم اما أن يخلق كل سلالة منفردة و الا فهو غير موجود و هكذا بمجرد أن يخضع أى كائن لأى تكيف مع بيئته فيتغير حجم منقار أو سمك صدفة أو عدد شعيرات ذبابة يعتبر هذا دليلا على حاسما على غياب التصميم فالمصمم يجب أن يخلق كائنات ان تغيرت بيئتها قليلا تسقط ميتة فورا و لا تتكيف معها و عليك الاكتفاء بالكلام الانشائى عن تراكم التكيفات عبر ملايين السنين و الا فانك تتهم المصمم بتضليل الباحثين الشرفاء عن الحق!
ناقشنا هذه المسخرة سابقا و وضحنا كيف أنه لا عقل و لا علم و لا واقع و لا حتى تاريخ البيولوجيا ذاته كما رواه رواد نظرية التطور فى القرن العشرين يقول بأن بالتصميم أو حتى الخلق يتعرض مع استخدام نفس المكونات و أن الانتظام الهرمى الذى يزعم التطوريون أنه دليل حاسم على التطور هو فى الواقع عملية تصنيف سابقة على النظرية سرقتها النظرية و نسبتها الى نفسها و لكن الحقيقة أن الأبحاث العلمية الحديثة كشفت عن المزيد. ماذا لو اكتشفنا أن هذه المكونات هى المكونات المثلى؟ أن كل مكون فى الواقع يوازن بين عدد كبير من الصفات المطلوبة لبناء أى شكل من أشكال الحياة؟ ماذا لو اكتشفت الأبحاث أن الكلام عن حياة من مكونات مختلفة كلام غير عملى لأن المكونات البديلة المقترحة دائما تحتوى على عيوب منع استخدامها فى أى عملية معلوماتية أو بيوكيميائية مطلوب منها أن تعيش و تتكاثر و تتكيف و تنتشر و تبقى على المدى البعيد؟ بل ماذا لو اكتشفنا أن المكونات الحالية هى الوحيدة الممكنة عمليا لدرجة أن "تطور"شكل مختلف من الحياة فى كوكب اخر يجب أن يستخدم مثل هذه المكونات؟ سيعتبر هذا دليلا اضافيا ضد "لاهوت"الصانع المخادع الذى كان يمكن أن يضع أى مكونات أخرى.
تحدثنا فى المقال السابق عن مثالية الأحماض الأمينية المشفرة فى الحمض النووى/الدنا DNA و الان نتحدث عن الحمض النووى ذاته. تتألف السلسلة الواحدة من الحمض النووى من مجموعة مكونات تسمى نيوكليوتيدات Adenine / Guanine / Cytosine / Thymine يمكن تشبيها بالحروف الأبجدية للغة و اختصاراتها A/G/C/T و تتصل هذه النيوكليوتيدات ببعضها البعض صانعة تسلسل من الحروف يشبه الكلمات و الجمل فى اللغة. تتألف النيوكليوتيدة الواحدة فى الدنا من جزئ من سكر الرايبوز ribose (للدقة رايبوز منزوع الأكسجين De-oxy) تتصل بها من جهة مجموعة فوسفات و من جهة أخرى قاعدة نيتروجينية متغيرة هى التى تمثل الحرف المطلوب A/G/C/T. [برغم كون الحرف ممثل فى القاعدة النيتروجينية تحديدا يتم استخدام مصطلح الحرف كثيرا للاشارة الى النيوكليوتيدة بالكامل]

صورة للثايمين T – مجموعة الفوسفات الى اليسار بالأصفر – حلقة السكر فى المنتصف الى الأسفل بالأزرق – القاعدة النيتروجينية الى اليمين فى الأعلى بالأزرق
انظر جيدا الى الترقيم الموجود فى الصورة خاصة الأرقام المكتوبة بالأحمر فى حلقة السكر (deoxy)ribose – هذا الترقيم هو ترقيم ذرات الكربون و العلامة الصغيرة بجوار الرقم (') و تنطق prime هى لتمييز أن الرقم ينتمى الى حلقة السكر و ليس الى القاعدة النيتروجينية. اتصال مكونات النيوكليوتيدة عند ذرات الكربون الصحيحة مهم لأنه يؤثر لاحقا على قدرة النيوكليوتيدات على أن تتصل معا فى شريط ذو مرونة/مدى حركى ملائم للطى و الانثناء و و قدرة الشريط المصنوع منها على الاتحام مع شريط مقابل لكن أكثر ما يهمنا الان هما الرقمان 3'/5' لأنهما حيث تتصل هذه النيوكليوتيدات معا لصناعة الشريط

لاحظ كيف تحدث الاتصالات بانتظام بين الحروف عن طريق مجموعة الهيدروكسيل OH عند 3' و مجموعة الفوسفات عند 5' صانعة ما يشبه السلم. هذا النمط فى الارتباط مهم للحفاظ على اتساق الهيكل و منع التفرعات الجانبية بحيث يصنع الرايبوز و مجموعات الفوسفات الألفة للماء ما يسمى بالعمود الفقرى للحمض النووى sugar-phosphate backbone بينما تمتد القواعد النيتروجينية فى الاتجاه المقابل تمهيدا لارتباطها بمثيلاتها فى الشريط المقابل

صورة تعبر عن بصيرة الانتخاب الطبيعى و التفاعلات الكيميائية – صنعت شريطا بهيكل و كأنه يعلم أن شريطا مقابلا سيرتبط به فى المستقبل
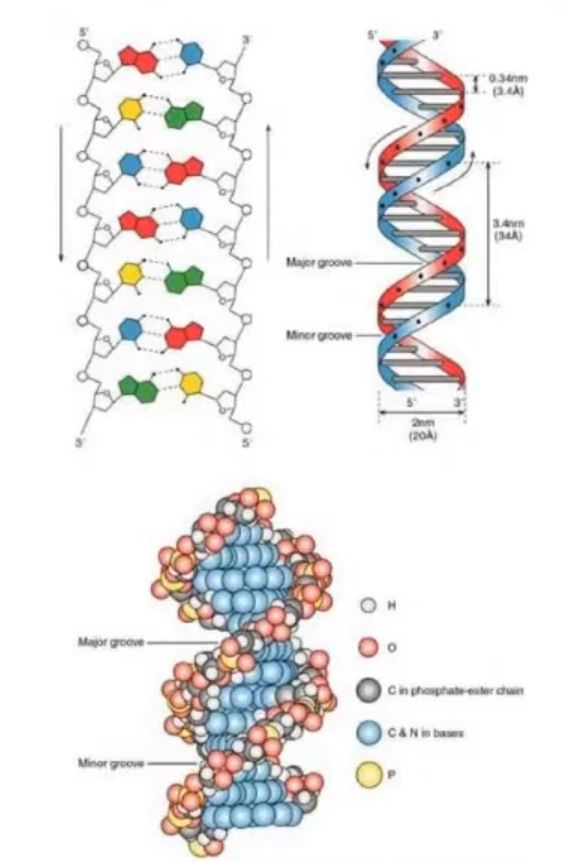
يتألف جزئ الحمض النووى من شريطين متقابلين و متعاكسين فى الاتجاه Anti-parallel يسير أحدهما فى اتجاه 3'الى 5' بينما يسير الاخر عكسه فى اتجاه 5' الى 3' و كأنهما طريقان متقابلان ذهابا و ايابا بحيث يصنع العمودان الفقاريان لكلا الشريطين الاطار الخارجى للجزئ بينما تلتقى القواعد النيتروجينية (درجات السلم) الكارهة للماء معا فى الداخل بحيث تتقابل A/T و C/G معا و تنشأ بين هذه الأزواج روابط هيدروجينية لتثبيت الهيكل.

لاحظ كيف أن كل شريط يكون مقلوبا/معكوسا عند مقارنته بالاخر ففى أحدهما فى حلقة الرايبوز من كل حرف (المظللة بالأزرق) ذرة الأكسجين O و ذرة الكربون الخامسة 5' بالأعلى بينما ذرة الكربون الثالثة 3' بالأسفل أما الشريط المقابل فيسير عكس الاتجاه بذرة الأكسجين و الكربون الخامس الى الأسفل و الكربون الثالث نحو الأعلى. هذا الاتجاه المقلوب للرايبوز/الشريط ذو أهمية وظيفية حيث أن انزيم النسخ polymerase يمكن تشبيهه بالسيارة التى تسير على الطريق و الاتجاه الذى تستطيع أن تسير فيه هو اتجاه الشريط و بالتالى لنسخ شريطين أحدهما مقلوب الاخر يجب اضافة خطوات مثل طى الشريط المقلوب جزءا جزءا ليساير الاخر أثناء النسح ثم لصق هذه الأجزاء المستقلة بعد انشاءها و كل هذا بالطبع يحتاج الى الات جزيئية و خطوات متناسقة (سنشرحها بالأسفل عند الحديث عن okazaki fragments قطع أوكازاكى)

لاحظ فى الصورة كيف أن الشريط الأسفل يدور أولا قبل أن يدخل فى الحلقة البنفسجية (انزيم النسخ) بحيث يدخل فيه معكوسا بينما يدخل فيه الشريط الأعلى مباشرة و فى المقطع الحلقة التى تنتفخ كالبالون الى اليسار ثم تنحل ثم يعاد عمل واحدة جديدة – هذه هى عملية طى/لف الشريط المعكوس – ينعكس المعكوس فيصبح متوازى و يسر كل مركب نسخ replisome كله فى اتجاه واحد
يدور هذا السلم بعد ذلك باتجاه اليمين لصناعة هيكل اللولب المزدوج double helix الشهير

هيكل السلم الملفوف يصنع مناطق مقعرة grooves كبيرة و صغيرة على طول الجزئ تصلح كمنصات هبوط و ارتباط للبروتينات التى ستعمل على الحمض النووى.
يمكن تقسيم القواعد النيتروجينية الأربعة الى نوعين البيورينات purines و هى الأدينين و الجوانين A/G و هذه أكبر حجما و تتألف من حلقتين bicyclic و البيريميدينات pyrimidines و هى السايتوزين و الثايمين(يوراسيل) C/T/U و هى أصغر حجما و تتألف من حلقة واحدة monocyclic

هذا هو الحمض النووى الرايبوزى منقوص الأكسجين و الذى يشتهر فقط بالحمض النووى/الدنا DeoxyriboNucleic Acid/DNA أما زميله الرنا RiboNucleic Acid/RNA فهو يشبهه فى المكونات و لكن الرايبوز ليس منزوع الأكسجين بل يحتوى على مجموعة هيدروكسيل OH

كما أن القاعدة النيتروجينية/الحرف thymine/t ليست موجودة و بدلا منها Uracil/U يوراسيل

الاستخدام الأشهر للرنا هو أدواره فى ترجمة الشفرة من الحمض النووى الى سلسلة أحماض أمينية و لكن ثبتت له استخدامات أخرى كثيرة فى الخلية
اذن فلدينا مكونات هيكلية أو أحجار بناء ان جاز التعبير متمثلة فى النيوكليوتيدات و مكوناتها من جهة و لدينا شفرة معلوماتية تستخدم لتخزين هياكل البروتينات (الكلمات و الجمل الوظيفية أو بشكل أدق الالات الجزيئية) المختلفة و طبعا كالعادة سيتحفك التطورى بالجملة التى لا يحفظ سواها و يكررها فى كل مجال من التشريح الى الكيمياء الحيوية الى علوم الجينات: ان هذه المكونات و الهياكل مشتركة بسبب وراثتها من سلف مشترك و بالتالى تتواتر الأدلة من هذه المجالات المختلفة على "حقيقة التطور"بل و ربما يزيدك التطورى قائلا أنه لا يوجد سبب للاشتراك فى المكونات غير السلف المشترك الا لو كان المصمم يخدعنا بل و ربما يحاول التجويد أكثر و أكثر زاعما أن هذه المكونات غير مثالية و يمكن استبدالها بما هو أحسن منها مما ينفى التصميم بالكلية و يرد على هؤلاء الخلقوين الذين يقحمون الدين فى العلم و يزعمون أن تشابه المكونات و الأسس بسبب ملاءمتها لوظيفتها. لو أنك قرأت مقالنا السابق عن الأحماض الأمينية فيمكنك أن تتوقع بسهولة ما ستجده هنا: نعم لقد أثبتت الكشوفات العلمية أن هذه المكونات من أحجار البناء الى الشفرة هى المثالية لأداء وظائفها و كما جرت العادة بمجرد سقوط تنبؤاتهم السابقة و ثبوت مثالية المكونات يقوم التطوريون باستحضار الانتخاب الطبيعى لانقاذ الموقف لينتقل صديقنا التطورى اللطيف الى مرحلة "أوه لقد أتى لنا الانتخاب الطبيعى بأفضل ما هو ممكن و وازن كل المتطلبات بشكل مثالى – ان السمكرى الأخرق مهندس رائع التصميمات"!
The similarity between the creations of tinkerer and engineer also raises a fundamental scientific challenge: understanding the laws of nature that unite evolved and designed systems.
U. Alon "Biological Networks: The Tinkerer as an Engineer" Science Vol 301, Issue 5641 pp. 1866-1867 (26 Sep 2003)
المشكلة الكبرى هنا هى أن صديقنا التطورى لا ينتبه (أو لا يريد أن ينتبه) الى عدة نقاط تأسيسية:
1- بمجرد أن نسب التطورى الشئ الى الانتخاب الطبيعى بسبب فوائده (من يملكه خير من من لا يملكه) و ملاءمته لوظيفة/غرض ما فقد انتهت تماما فكرة عدم امكان تفسير تشابه المكون الا بالسلف المشترك أو فكرة المصمم المخادع فالمصمم بداهة لا يخدعك لأنه يضع المكون المناسب بل انك و يا للعجب ستجد تيمة "لماذا اختارت الطبيعة كذا و كذا" متكررة فى أوراقهم و هى تيمة تتعارض تماما مع فكرة عدم خصوصية المكونات و لو صاغ لهم أحد السؤال بصيغة "لماذا اختار المصمم" لاتهموه بمحاربة العلم! ليس ذلك فحسب بل ان بعض التفسيرات التطورية قد تتجاوز فكرة الانتخاب الى ما يسمى بالحتمية الكيميائية – بعبارة أخرى ما كان مكون لينجح هنا أو حتى فى كوكب اخر سوى هذا و بالتالى لا حاجة لمصمم لينتقى أى شئ و هذا فى الواقع اعتراف أكبر بمثالية و خصوصية المكونات و بسقوط فكرة الصانع المخادع الذى يوحى لنا بالسلف المشترك بل هو الصانع الذى صنع لكل وظيفة مكون مثالى
2- الخيال العلمى يختلف عن العلم! افتراض أن مجرد وجود المكون المثالى معناه أن التطور سيصنع منه شيئا مبنى على المبالغة بشدة فى قدرات الانتخاب الطبيعى و لقد شرحنا من قبل عجزه عن حمايتنا من التدهور الجينى بل و قيامه بانتخابه أحيانا و لقد رأينا فى مقال الأحماض الأمينية كيف يطلق التطوريون مزاعم عن قيامه بتعديل الشفرة و المكونات بلا دليل لمجرد سقوط تنبؤاتهم السابقة بسوء تصميمها ثم يعودون هم للاعتراف بكم التدخل "العلمى/الذكى" المطلوب للقيام بتعديلات أقل بكثير. أضف الى ذلك أن الانتخاب يتم استخدامه كثيرا كعملية تعمية/قنبلة دخان تحجب التساؤلات الحقيقى: هل فعلا هناك مجال لتوليد ما يكفى من التباينات و هل فعلا هناك خطوات وظيفية نحو كل شئ و هما سؤالان اجابتهما فى معظم الأحوال: لا مهما حاول التطورى تجاوز الحسابات بكلمات: ملايين السنين/خطوة خطوة
3- حتى اذا أردت تجاهل النقطة السابقة و افتراض قدرات كبيرة للانتخاب الطبيعى و خطوات وظيفية مفترضة فان هذه المكونات موجودة منذ اللحظة الأولى فى أبسط الكائنات الحية و أقربها تطوريا للسلف المشترك العالمى المفترض بل كما بيننا فى المقال السابق الذى يوضح مثالية الأحماض الأمينية يرجحون وجودها فيه هو ذاته و بالتالى لا مجال للانتخاب الطبيعى ليحسن أى شئ مما يضطر التطوريون الى افتراض شفرات سابقة على الشفرة الحالية و عوالم الرنا RNA world و ما قبل الرنا pre RNA world و كل هذه فرضيات خيالية و عندما تضيف الى ذلك أنهم يرجحون ظهوره هذا السلف المشابه للبكتيريا المعاصرة بعد القصف النيزكى أو أثناؤه منذ أكثر من 4 مليار سنة لا يتبقى أى مجال ليتطور أى شئ حتى لو أردت أن تفترض امكان حدوث ذلك التطور ابتداءا
4-اذا فشلت الاعتراضات السابقة يلقى اليك التطورى بورقة الدائرة المربعة ليعود بك صراحة الى مربع اللاهوت (الذى لم يغادره فى الواقع) و الى الحديث عن قدرات المصمم و ما يتوقعه هو منه. لماذا ينتقى المصمم مكونات وفق قواعد و قوانين و هو مطلق القدرة قادر على صناعة ما يريد بدون هذه القواعد؟ بعبارة أخرى: أين الدائرة المربعة؟ طالما هى غير موجودة و كل شئ يسير وفق قواعد الطبيعة فلا يوجد تصميم ليقوم التطورى بعملية التمويه المفضلة لديه والتى قام بها دوبزانسكى: تجاوز عدم وجود أى الية طبيعية لانتاج ما أمامنا و اعتبار التشغيب اللاهوتى على فرضية التصميم يدل ضمنيا على وجود هذه الالية. فقط تخيل نفسك تقول هذا الكلام لمهندس و أنتما تنظران الى المبنى الذى صممه: بما أنه لا توجد دوائر مربعة فى المبنى فأنت لم تصممه! و قبل أن يتحفنا التطورى بالفروق بين حدود قدرة مصمم المبنى و مصمم الكون دعنا نذكره بأن زعمه الأصلى كان أن تشابه المكونات لا يمكن تفسيره بالتصميم خاصة و أن المكونات لا تحمل خصوصية فلما اكتشف أن المصمم عظم قيمة العلم و العقل و ترك له فى خصوصية المكونات بصمة يراها بعينيه و يرصدها بأدواته و يدركها بعقله ليثبت له التصميم مسقطا زعمه - عكس كلامه و أصبح يطالب المصمم بصناعة كائنات من مكونات لا تحمل أى خصوصية بل و ربما صناعتها بما يشبه السحر الذى لا يمكن دراسته علميا!!! نفس التطورى ان وجد أمامه كائن حى من أى مكونات غير مثالية لهلل فرحا و اعتبر ذلك دليلا على عشوائية الطبيعة و عملها كسمكرى يصنع الكائن من اى ترقيعة من المواد حتى لو لم تكن الأفضل – بعبارة أخرى التطورى كان يريد من المصمم صناعة كائنات توحى بالسمكرة و التطور أو على أفضل تقدير يتم تصنيفها كظواهر عصية على الفهم و الدراسة المعملية و بالتالى لا يمكن الاستدلال بها علميا و هذا اعتراض من نوعية اما أن أربح أنا أو أن تخسر أنت!
5-هذا المقال يتعامل مع المرحلة التالية لفرضيات انتاج مكونات الحياة فى بيئة الأرض البدائية و السابقة للخلية الأولى و لكن ليس معنى ذلك التسليم للتطوريين بما يقولوه فى هذه الفرضيات فما يفعلونه فى الواقع هو بناء طبقات من الفرضيات و القصص بعضها فوق بعض و عند نقاش أى فرضية يتعاملون و كأن ما قبلها و ما بعدها صحيح و هو ليس كذلك. لنا هنا على المدونة مقالات سابقة تتحدث عن استحالة نشأة مكونات الحياة و بقاءها دون اتلاف فى بيئة الأرض البدائية و استحالة تفاعلها معا بالشكل المطلوب ان بقيت دون تداخل من المكونات الأخرى أو من طبيعة المكونات نفسها و كمية التدخلات "المعلوماتية"التى يقومون بها فى "تصميم"التجارب و عمليات المحاكاة حتى يتم تركيز المكونات و انتظامها فى الهياكل و التسلسلات المطلوبة و تبقى دون تلف ثم يفترضون أن الطبيعة ستقوم بالمثل لمجرد أن الأرض نظام مفتوح ناهيك عن طبيعة شكل الحياة الأبسط القادر على التواجد مستقلا من واقع الطبيعة و ليس من قصص الناسخ الذاتى الأول الخيالية أو من توسلات على شاكلة "لابد أنها كانت كذلك و الا فلن يعمل التطور"
العاملون فى المجال يتجاهلون كل هذا و يقفزون مباشرة الى تصميم رايبوزايم ribozyme (شريط متطوى/انزيم من الرنا) فى المعمل مفترضين أن التفاعلات الكيميائية ستحاكى ما فعلوه بالعلم و أن الطبيعة مثل بيئة المعمل المصممة بعناية لحمايته و منعه من التلف و التفاعلات الجانبية من جهة و امداداه بشحنات اضافية من المكونات التى يحتاجها فى النسخ و التكاثر من جهة و مفترضين أنه بمجرد وجود الرايبوزايم فهناك مسار ما ليتحول الى خلية فى النهاية لذا عندما نتحدث عن الناسخ الذاتى الأول فى هذا المقال فى سياق مناقشة مكونات الحمض النووى/الرنا فهذا ليس تسليما بفرضياتهم حول بيئة الأرض و التفاعلات و شكل الحياة الأول و قدرة الطبيعة على محاكاة المعمل بل هو لتبيان ما هو مضمن فى هذه الطبقة من الفرضيات و كيف تتسق البيانات أكثر مع التصميم و أمثلية المكونات مما يسقط تماما تفسيرها بالسلف المشترك. و أخيرا تذكر أنه وفقا لفرضياتهم فان كل المكونات المثالية بعد أن نشأت فى بيئة الأرض البدائية و تفاعلت مع غيرها من المكونات المثالية حصرا لانتاج الحياة قام التطور بانشاء مسارات بيوكيميائية لانتاجها هى تحديدا دونا عن أى شئ اخر و هذه بمفردها كارثة لمن يفهم تعقيد المسارات البيوكيميائية و خطواتها المتعددة و الانزيمات المتخصصة التى تقوم بها وهو أمر يحتاج مقالات متعددة لشرحه و هذا الكلام ليس قاصرا على الحمض النووى و الرنا اللذان نتحدث عنهما هنا بل على الأحماض الأمينية و كل مكون اخر يتم تصنيعه فى الخلية

ملحوظة: سترد مصطلحات مثل شريط/منفرد/وحيد عند الحديث عن الرايبوزايم و معناها فى السياق هو غياب الالتحام مع قالب مقابل و ليس بالضرورة غياب القالب المقابل ذاته بالكلية لأن الالتحام سيغلق الشريط أمام عملية النسخ و سيحتاج الات متخصصة لفتحه و التعامل معه (انظرالصورة/المقطع الخاصين بقطع أوكازاكى) و هو ما لا يفترضون وجوده فى بيئة الأرض الأولى
هذه النقاط الخمسة أبقها فى رأسك باستمرار و بالذات النقطة الأولى لأن الهدف الأساسى للمقال هو التعرض لفرضيات التنادد homology التى تزعم أن تشابه مكونات الكائنات الحية من المصمم خداع غير مفهوم و ظاهرة لا تفسر الا بالوراثة من سلف مشترك! لاحظ أيضا كيف يتحول سؤال "لماذا"الى محرك للاكتشاف و دافع للعلم عندما يتم سؤاله بغرض البحث عن اجابة فعلا و ليس بغرض الاستهجان على طريقة "لماذا المكونات متشابهة – لماذا الأطراف متشابهة – لماذا الشفرة متشابهة هل يخدعنا المصمم ليوحى لنا بالتطور من سلف مشترك؟"
لماذا الفوسفات؟
سنة 2010 صدرت ورقة علمية حول اكتشاف شكل من أشكال الحياة يمكنه الاعتماد على الأرسينيك(زرنيخ) بدلا من الفوسفور و أثارت جدلا واسعا حول مدى امكان صحة هذا الاستنتاج الى أن تم سحبها بعد 15عاما سنة 2025.
"Controversial ‘arsenic life’ paper retracted after 15 years - but authors fight back" Nature News (24 July 2025)
"Fifteen years later, Science retracts 'arsenic life' paper despite study authors" Science News (24 July 2025)
"'Arsenic Life' Microbe Study Retracted after 15 years of controversy" Scientific American (24 July 2025)
أحد الاسباب التى جعلت هذه الورقة تتعرض الى انتقادات كثيرة هو ادراك الكثير من العلماء أن استبدال مكون من مكونات الحياة باخر عمليا مستحيل برغم سيناريوهات الخيال العلمى النظرية التى يطلقها التطوريون يمينا و يسارا اذ يقول كثير منهم أن هذه المكونات قد جاءت عن بلا تخطيط أو تقدير أو تصميم مسبق و كان يمكن ابدالها بأخرى و بالتالى فانتشارها بين الكائنات لا سبب له سوى الوراثة من سلف مشترك – على سبيل المثال تقول بعض الفرضيات التطورية أنه وسط فوضى التفاعلات العشوائية فى بيئة الأرض القديمة كان حظنا أن نشأت سلسلة الناسخ الذاتى الأول معتمدة على مجموعات الفوسفات كأحد المكونات ثم "طورت"خصائص متعددة فى مجال توليد و تخزين الطاقة و صناعة غلاف خارجى أيضا باستخدام مكونات تعتمد على الفوسفات و كان من الممكن أن تنشأ هذه الوظائف باستخدام كيمياء أخرى لنكتشف لاحقا أن المسألة لم تكن مصادفة فى الحساء البدائى بل أن مجموعة الفوسفات هى الأمثل للوظائف المطلوبة فمثلا منذ بداية دراسة مكونات الرنا و الحمض النووى جاءت الأبحاث مؤكدة أن صفات الفوسفات مثل نوعية روابطه و شحنته السالبة و موقعها فى هيكل الجزئ هى الأمثل للوظيفة المطلوبة بعد مقارنتها بالبدائل المتاحة و التى كان منها الأرسينيك ذاته و الذى طار به البعض كدليل على امكان تنوع مكونات الحياة و عدم خصوصيتها و بالتبعية صدفويتها
No other residue appears to fulfill the multiple roles of phosphate in biochemistry… In summary, the existence of a genetic material such as DNA requires a compound for a connecting link that is at least divalent. In order that the resulting material remain within a membrane, it should always be charged, and therefore the linking unit should have a third, ionizable group. The linkage is conveniently made by ester bonds, but, in order that the ester be hydrolytically stable, that charge should be negative and should be physically close to the ester groups. All of these conditions are met by phosphoric acid, and no alternative is obvious. Furthermore, phosphoric acid can form monoesters of organic compounds that can decompose by a mechanism other than normal nucleophilic attack, a mechanism that allows them sufficient reactivity to function in intermediary metabolism. Finally, we can answer the question concerning the choices made by chemists and by natural selection...We can understand the choices made both by chemists and by the process of natural selection. They are both correct.
F.H. Westheimer “Why nature chose phosphates?” Science 235: 1173-78
الطريف فى الموضوع أن الباحث يقارن "قرارات" الانتخاب الطبيعى بقرارات العلماء موضحا فرق سياقات التجارب عن سياقات الخلية و بالتبعية عدم امكان الاستدلال بما هو صالح لأحدهما على الاخر فقارن العقل و العلم مع الطبيعة العمياء و لو قال له أحدهم قرارات "المصمم"لاتهمه بالخروج من دائرة العلم! الأطرف أن هذه المكونات لا يمكن نسبتها للانتخاب الطبيعى لأنها يجب أن تنشأ مع الناسخ الذاتى الأول من الكيمياء العشوائية فى بيئة الأرض القديمة فيجب أن تكون صحيحة من الضربة الأولى. الأهم من ذلك أنه يشير الى بعض البدائل التى قد يكون عمرها قصيرا و هى بدائل صالحة لناسخ ذاتى بدائى أو حتى للبكتيريا و لكنها غير صالحة للكائنات الأطول عمرا! بعبارة أخرى يبدو أن الانتخاب الطبيعى كانت لديه رؤية مستقبلية لما سيحدث بعد الناسخ الذاتى الأول!!!
If a single ester bond in the genetic material is cleaved during the lifetime of the organism, it may fail to reproduce. Of course, with bacteria, a small sample may contain many billions of organisms, and only a few of them need to reproduce to maintain the species. Still, natural selection will favor a genetic material that, in the majority of the cells, will last for times comparable to the lifetime of the organism. Certainly higher organisms, whose numbers are limited, would have special difficulties with labile genetic material…esters of carboxylic acids would be marginal as a genetic material for rapidly dividing bacteria and totally inadequate for longer lived animals or plants. Some sterically hindered esters that are more stable might serve, but the system would become unnecessarily complicated.
و منذ ذلك الحين توالت الأبحاث التى تستحسن اختيار "الطبيعة" للفوسفات فى وظائفه المتعددة من الحمض النووى الى الرنا الى دهون الغشاء phospholipids الى عملة تبادل الطاقة فى الخلية الأدينوسين ثلاثى الفوسفات ATP. سنفرد لهذه الأشياء مقالة مستقلة ان شاء الله لكن نكتفى الان بتنبيه القارئ الى هذه المفارقة و ان كانت ليست موضوعنا الأساسى: أن يكون مكونا ما – بالصدفة – هو الأفضل ملاءمة لعمليات مختلفة مطلوبة للحياة و كأن "الطبيعة" نحتت هذا المكون خصيصا لاستخدامه كحجر من أحجار بناء الحياة.
proteins exploit the charge shift upon motion from the reactant to TS[transition state] in phosphate hydrolysis, to convert a very high-activation barrier in solution to a relatively low activation barrier, and thus gain complete kinetic control on many biological processes (see e.g. the discussion of ATP hydrolysis in Section 4). Next, the electrostatic repulsion between negatively charged phosphate fragments is used as a source of biological energy (again a point that was not clearly analyzed before). Finally, having negatively charged phosphates and changing the magnitude of the negative charge during the reaction (e.g. GTP being hydrolyzed to GDP) provides an ideal way for allosteric control of protein function and signal transduction (see Section 4.5).
Shina C L Kamerlin et al., "Why nature really chose phosphate" Quarterly Reviews of Biophysics Volume 46 Issue 1 (2013) p. 1-132
لدرجة أن الملحد الشهير بيتر أتكنز وصفه بأنه العنصر المثالى لنقل الطاقة فى الخلايا
Phosphorus, in the form of adenosine triphosphate (ATP) turns out to be a perfect vector for the subtle deployment of energy, and it is common to all living cells.
Peter Atkins, The Periodic Kingdom. p. 27
و شرح علماء اخرون مثالية مقدار الطاقة فى روابطه لعمليات الخلية من جهة و كونها ليست سهلة التفاعل من جهة أخرى
[the amount of energy released on hydrolysis of the terminal phosphate] is comparable to the energy consumed in many kinds of biochemical transformations
Rob Phillips et al., “Physical Biology of the Cell” 2nd edition (2013) p.192
ATP itself is relatively stable and hence is unlikely to adenylate or phosphorylate other molecules in the absence of suitable catalysts
Silvana Pinna et al., “A Prebiotic Basis for ATP as the Universal Energy Currency, PLoS Biology 20 (October 4, 2022): e3001437

رابط الفوسفات الثلاثى – الرابط المثالى كعملة تحزين و استهلاك طاقة
لاحظ أن الأبحاث تقر بوجود مركبات أخرى قد تقوم بأدوار كثيرة مشابهة للفوسفات و لكن عادة ما تكون هناك صفة أو أخرى تجعلها غير ملائمة فى سياق الخلية مما يطعن بشدة فى أسلوب البعض بذكر خصائص مركبات شبيهة بالفوسفات و الزعم بأنها بدائل ممكنة. لاحظ أيضا كيف أن بعض السمات التى اختارت من أجلها "الطبيعة" الفوسفات صفات مستقبلية تحتاج الى بصيرة ككون الانزيمات المتعاملة معه أقل تعقيدا من المطلوبة للبدائل و كأن الطبيعة كانت تعلم أنها ستتطور انزيمات فى المستقبل!
the cleavage of phosphorus-oxygen bonds is arguably the most important biological reaction, being involved in everything, from signaling and energy transduction to protein synthesis and maintaining the integrity of the genetic material...catalyzing phosphoryl transfer is notoriously demanding, with half-lives for the cleavage of phosphorus-oxygen bonds extending into millions of years...This immediately gives phosphate esters an advantage as a biologically relevant compound, as the existence of life as we know it depends on the tight regulation of interrelated chemical reactions occurring in vivo, where a simple difference in rate of a factor of 2 can be the difference between life and death. Such tight regulation is impossible if the reactions being regulated could occur spontaneously, which is why we are also highly skeptical of the idea that arsenate can replace phosphate in biological systems...despite the tremendous rate accelerations enzymes need to achieve to cleave phosphorus-oxygen bonds, phosphate esters are highly versatile...they can proceed through a multitude of different mechanisms, and also, equally importantly, can act as their own proton sink for nucleophile activation, giving them an immediate evolutionary advantage over, for instance, sulfuryl transfer reactions...This is doubly important in light of the fact that sulfates fulfill many of the same biological functions as phosphates; however, it seems to appear that they are even more inert than phosphate esters in aqueous solution (it has been suggested that S-O cleaving enzymes as the most proficient known to date, Edwards et al. 2012), and, the requirement of an ‘external’ base makes it more challenging to evolve for sulfatase activity than for phosphatase activity, which perhaps explains the prevalence of phosphate over sulfate esters in biology.
Shina C L Kamerlin et al., "Why nature really chose phosphate" Quarterly Reviews of Biophysics Volume 46 Issue 1 (2013) p. 1-132
ملحوظة: ستكون لنا ان شاء الله وقفات مطولة أخرى مع الفوسفات سواء كمكون لدهون غشاء الخلية أو كعملة نقل الطاقة ناهيك عن مناقشة الدورات الاستقلابية و مسارات توليد الطاقة فى الخلية Glycolysis/krebs cycle/Electron transport chain لتوضيح خطأ مزاعم التطوريين بشأن كونها عمليات غير مثالية و قابلة للتحسين و مكوناتها قابلة للاستبدال و لاثبات كونها مصممة بعناية ومثالية لأهدافها ردا على مزاعم التطوريين أن التفسير الأفضل لوجودها هو السلف المشترك و لكن ما سنركز عليه الان هو الحمض النووى تحديدا ردا على مزاعم أنه دليل على التطور و سننتقل الى المكون الثانى
لماذا الرايبوز ribose؟
كالعادة ليست المسألة أن مكونا ما من بيئة الأرض البدائية قد دخل فى التفاعلات لينتج حجر بناء كان يمكن استبداله باخر بل تشير الأبحاث الى مثالية السكريات خماسية الكربون pentose كمكون للنيوكليوتيدات و من بين أنواعها مثالية سكر الرايبوز تحديدا للوظيفة المطلوبة و ليس أى رايبوز لأن المركب الكيميائى يأخذ شكل حلقة و الحلقات بدورها يمكن أن تأخذ عدة أشكال على حسب هل الحلقة خماسية (مع دخول ذرة أكسجين فى أحد الأركان و بقاء ذرة كربون خارجها)furanose rings أو سداسية pyranose تدخل فيها ذرة الكربون الباقية و هل ترتفع مجموعة الهيدروكسيل عند ذرة الكربون الأولى الى الأعلى (ألفا) أم تنخفض الى الأسفل (بيتا)

ملحوظة: الصورة تظهر تنوعات الرايبوز فقط و لا تظهر باقى السكريات الخماسية بتنوعاتها ناهيك عن السكريات الغير خماسية بتنوعاتها وسط طوفان المكونات الأخرى فى بيئة الأرض البدائية المفترضة و من بين كل هذا يتضح لنا أن حلقة فيورانوز الخماسية للبنتوز رايبوز بيتا-دى beta-d تحديدا المستخدمة فى حمضنا النووى هى الأمثل لهذه الوظيفة لما توازنه من مواصفات متعددة كالحجم و الهيكل و المرونة الحركية puckering
the five-carbon sugars used in nucleic acids are likely to be repeated themes, perhaps in part because they are the smallest sugars that can cyclize and thereby confer spatial orientation on other molecules, for instance the purines and pyrimidines that comprise the genetic information of terrestrial organisms.
Pace, N.R. (2001) The universal nature of biochemistry. Proc Natl Acad Sci USA 98:805–808.
Consequently, [alpha]-nucleotides could not be selected as sugar components of nucleic acids. This notion is confirmed by the biosynthesis of purine nucleotides starting the assembly of purine ring on [alpha]-D-ribose. The configuration of ribose at C1' is inverted to [beta] in the second step of biosynthesis when the amide group of glutamine displaces the pyrophosphate in 5'-phosphoribosyl-1'-pyrophosphate (PRPP). Similarly, the pyrimidine biosynthesis also involves PRPP where ribose is initially an [alpha]-anomer. The [alpha] configuration of ribose is inverted to [beta] during the formation of orotidylate from orotate
Gaspar Banfalvi “Why Ribose was selected as the sugar component of Nucleic Acids” DNA and cell Biology vol. 25, no. 3, pp. 189–196.
the selection of ribose as the exclusive sugar component, as only this pentose fits perfectly into the known nucleotides...in all naturally occurring biomolecules containing D-ribose, it is present in the form of [beta]-D-ribofuranose...Among the structural constraints, the stability, ability to form complementary base pairs, linear covalent polymers, the encoded information, compaction in small spaces such as cells, could be mentioned.
only [beta]-D-ribose could be inserted into nucleotides to secure the free rotation for the functional groups (OH, phosphate, and base) and the stability, including the possibility of forming a double-stranded structure. Unrestricted rotation of functional groups in ribonucleotides is possible only with [beta]-D-ribose. The conformers originating from sugar pucker folding as well as the pentose configurations indicated that the selection of ribose was not a random, but the only possible solution as only [beta]-D-ribose is fitting into a functional ribonucleic acid. By trying to fit any other pentose into the nucleotide, a sterical barrier is created...The advantage of selecting [beta]-D-ribose over [alpha]-D-ribose was that, in nucleotides, the C1'-base and C2'-OH substituents of ribose became localized distantly enough to move freely and provide flexibility
Gaspar Banfalvi "Ribose Selected as Precursor to Life" DNA AND CELL BIOLOGY Volume 39, Number 2, 2020 p. 177-186
كما ترى مما سبق بعض السمات التى جعلت "الطبيعة" تختار الرايبوز صفات مستقبلية لن تظهر أهميتها لدى الناسخ الذاتى الأول الذى يفترض كونه شريط من الرنا فهى صفات متعلقة بامكان تشكيل الحمض النووى لهيكل اللولب المزدوج بأزواج التيوكليوتيدات المتقابلة الذى لن يظهر-فى القصة التطورية-الا لاحقا
Conformations of Watson-Crick type polynucleotides are restricted to torsion angles with nucleotides in their most preferred “rigid” forms…In [alpha]-nucleotides sugar puckering can be regarded as the mirror image of[beta]-nucleotides...bases in [alpha]-anomeric position are unable to base pair, eliminating the possibility of helix formation.
Gaspar Banfalvi “Why Ribose was selected as the sugar component of Nucleic Acids” DNA and cell Biology vol. 25, no. 3, pp. 189–196.
بل و ربما كان الانتخاب الطبيعى (الذى لا يعمل قبل نشأة الناسخ الذاتى أصلا) قد اختار الرايبوز كمكون للناسخ الذاتى بناءا على ملاءمته لعمل انزيمات ستظهر لاحقا أثناء التطور مثل Rnase و ribonucleotide reductase لدرجة أن مكونات ملائمة بديلة مثل أرابينوز ربما لم يتم اختياره بسبب عدم ملاءمتها لعملية صناعة الحمض النووى التى لن تظهر الا فى المستقبل
functional groups attached to the furanose ring must have played an important role when ribose was selected as the sugar component of nucleic acids. This notion has been confirmed by synthetic nucleotides with modified sugars, which influence sugar puckering, and when incorporated into RNA significantly contribute to its RNase resistance.
The removal of 2'-OH in 2'-deoxyarabinose would solve the problem posed by the steric hyndrance. Eventually 2'-deoxyarabinose is equivalent to 2'-deoxyribose, and could be used as a building block in DNA but not in RNA since deoxyribonucleotides are formed from ribonucleotides...In arabinonucleotides, due to the presence of the bulky base at C1', access to the 2'-OH would be denied for the ribonucleotide reductase enzyme
يظهر لنا هنا كيف أن القاء كلمة الانتخاب الطبيعى يمينا و يسارا كلام بلا أى معنى حقيقى فهو لن يعرف أنه مستقبلا سيحتاج الى صناعة حمض نووى رايبوزى منقوص الأكسجين و سيحتاج الى انزيم ليصنع ذلك و يحب أن يتمكن الانزيم من الارتباط بالمكونات و لن يعرف أنه سيحتاج الى لولب مزدوج فيجب أن تلاءمه المكونات و لن يعرف أنه سيحتاج الى انزيمات تحطيم الرنا و يجب عليه أن ينتقى مكون يمكن للانزيم المستقبلى من الارتباط به!
و انزيم ribonucleotide reductase هو الذى يقوم بازالة جزيئة الأكسجين لتحويل الحمض النووى الرايبوزى RNA الى الحمض النووى الرايبوزى منقوص الأكسجين DNA و لهذا حكمة لا علاقة لها بمجرد تصادف وجود المكونات فى الحساء البدائى أو تطور انزيم بالصدفة ليقوم بتفاعل ما فبقاء ذرة الأكسجين المرتبطة بذرة الكربون الثانية فى الرايبوز (مجموعة الهيدروكسيل) هو ما يجعل الرنا سريع التحلل و بالتالى ملائم لوظيفة الذاكرة المؤقتة RAM التى يقوم بها فى الخلية اذ أن بقاءه فترة طويلة معناه استمرار انتاج البروتينات لفترة أطول من المطلوبة بينما كونه غير مستقر و أسرع من الدنا فى التحلل الذاتى يخفف العبء عن اليات تحطيمه فى الخلية
Hydrolysis of RNA. Although RNA is a phosphodiester and carries a negative charge, it is relatively susceptible to hydrolysis; the rate of its spontaneous reaction with water, extrapolated to room temperature, is about 100 times greater than that of DNA. This occurs because the 2'-hydroxyl group of RNA acts as an internal nucleophile and leads to the formation of a 2',3'-cyclic phosphate with cleavage of the RNA chain. Intramolecular reactions are generally enormously more rapid than the corresponding external ones. In this case, the cyclic phosphate that is formed is highly strained, but the cyclization proceeds nonetheless, presumably because of the large positive entropy of the cleavage reaction. The cyclic phosphate in turn readily undergoes hydrolysis
F.H. Westheimer “Why nature chose phosphates?” Science 235: 1173-78

طبعا كعادة التطوريين فهم يتعاملون بطريقة بما أنه أنسب للوظيفة اذن فلابد أن التطور صنعه و هذا كلام فارغ و هذه السمات ذاتها التى تجعل الرنا ملائما لوظيفته تجعله غير ملائم لسيناريوهات عالم الرنا و الناسخ الذاتى الأول المزعوم و حتى الاستدلال بفيروسات الرنا هو استدلال بنموذج تنطبق عليه ظروف مختلفة تماما اذ حوله كبسولة حماية و لديه ميزة عددية هائلة لا يملكها ناسخ ذاتى يتيم نشأ بالصدفة من تفاعل كيميائى (بفرض امكان هذا السيناريو تنزلا) و لا يمتلك أى حماية
Since RNA is hydrolyzed relatively easily, it is not as well adapted as is DNA as a storage material for genetic material. The evolutionary advantage of DNA is apparent, although the evolutionary pathway to the difficult reduction process is obscure. Some viruses contain RNA and not DNA. However, in the tight viral package, water is excluded, so that the hydrolytic instability of RNA is not relevant. Subsequently, the time for viral reproduction in a cell during which the viral RNA is exposed to water is only a few minutes. In addition, not all of the enormous number of viral particles need reproduce to preserve the organism.
F.H. Westheimer “Why nature chose phosphates?” Science 235: 1173-78
ليس هذا فحسب بل ان مجموعة الهيدروكسيل هذه مهمة لقيام الرنا بوظائفه التحفيزية و التفاعلية و هى وظائف طبعا غير مطلوبة للدنا (طبعا من نافلة القول أن نوضح أن وجود وظيفة للشئ لا تجعله سلفا مشتركا و لن تجعله ينشأ من تفاعلات كيميائية عشوائية)
The oxygen atom that is removed by the reductase is of crucial importance to many ribozyme functions, since the 2'-OH is a strong nucleophile that forms transitional states during catalysis...substitution of NTPs by dNTPs on the active site of ribozymes would inactivate catalysis
Pedro E. Leon "Inhibition of Ribozymes by Deoxyribonucleotides and the Origin of DNA" Journal of molecular evolution (1998) 47:122–126
فى المقابل فان ازالة ذرة الأكسجين هذه و تحويله الى رايبوز منقوص الأكسجين DNAيجعل الجزئ أكثر استقرار و بالتبعية أنسب لتخزين المعلومات لفترات طويلة (وظيفة الجينوم)
From a chemical point of view the difference between RNA and DNA is rather small: a reduction of the 2'-hydroxyl group on the ribose moiety and a methylation of U to yield T (5 -methyluracil). The biological consequences of these changes are, however, considerable. The removal of the 2'hydroxyl group has been shown to chemically stabilize the genetic message by making the phosphodiester backbone less susceptible to hydrolysis and damage caused by UV irradiation.
Jorgen Jonsson et al., “The evolutionary transition from uracil to thymine balances the genetic code” Journal of Chemometrics 10, no.2(1996): 163
و يقلل من المساحة المشغولة فى قلب اللولب المزدوج ليتسع للقواعد النيتروجينية المتقابلة (النيوكليوتيدات/حروف الشفرة) مع اضفاء مرونة حركية pucker على حلقة السكر تساعد على تشكلها conformation فى الفراغ ثلاثى الأبعاد بشكل يجعل النيوكليوتيدات فى الحمض النووى قادرة على الارتباط بمثيلاتها على الشريط المقابل بطريقة تسمى hoogsteen/hg غير الطريقة التقليدية المسماة على اسم واطسون و كريك WC (يمكن محاولة تبسيطها بأن تخيل أن الجزئ يدور حول نفسه ليقوم بعمل الرابط بجانبه الاخر)

An HG base pair is created by rotation of a WC purine base ∼180° around the glycosidic bond, so that it adopts a syn rather than anti conformation (Fig. 1b). The two bases are also brought ∼2.0–2.5 Å closer, thus allowing formation of a unique set of hydrogen bonds (Fig. 1b). HG base pairs exist transiently (with typical lifetimes of 0.1–1 ms) and in low abundance (with populations typically <3%) in naked canonical B-DNA. However, dA-dT and dG-dC+ HG base pairs can become the dominant configuration in DNA–protein and DNA–small molecule complexes, in which these base pairs contribute to DNA recognition; in damaged nucleotides, in which they contribute to damage accommodation and repair; and in the active sites of translesion synthesis polymerases that use HG pairing to bypass damage during DNA replication
Huiqing Zhou et al., "m1A and m1G Potently Disrupt A-RNA Structure through the Intrinsic Instability of Hoogsteen Base Pairs" Nature Structural and Molecular Biology 23(9): 803–810.(2016)
Gaspar Banfalvi “Why Ribose was selected as the sugar component of Nucleic Acids” DNA and cell Biology vol. 25, no. 3, pp. 189–196.
هذه الطريقة تساعد على تحمل أى اضطرابات فى الهيكل مثلا أثناء فتح و اعادة طى اللولب المزدوج أو أثناء ارتباط البروتينات به أو تعرضه لضغوط أخرى كحدوث تلف مثلا فى موضع معين بينما فى الرنا بدون هذه المرونة الحركية يضطرب الهيكل و قد ينهار meltان حاول تكوين هذه الروابط. هذه الروابط بالتبعية مهمة لوظائف الحمض النووى فى تخزين و حفظ المعلومات و التعامل المستمر مع بروتينات الفك و الطى و القراءة و تحمل التلفيات المحتملة لتحقيق هدف التخزين طويل المدى للمعلومات
Duplex B-DNA can stably accommodate dA-dT and dG-dC+ HG base pairs, which can in turn play roles in sequence-specific DNA recognition, damage induction and repair, and DNA replication. In contrast, our results indicate that rA-rU and rG-rC+ HG base pairs are so unstable in the more compressed A-RNA that melting is preferred over the HG base-pair conformation.
The markedly different stability of the A-T/U and G-C+ HG base pairs in RNA and DNA duplexes provides a basis for achieving opposing functions at the genome and transcriptome levels (Fig. 5). If DNA did not have a capacity to form HG base pairs, and instead behaved similarly to RNA, lesions, such as m1dA and m1dG, that block canonical WC base pairing could greatly destabilize the double helix and potentially cause genomic instability (Fig. 5). The ability to form HG base pairs therefore endows DNA with an additional layer of chemical stability than is present in its RNA counterpart; this stability goes beyond resistance to hydrolysis, owing to the absence of the sugar 2′-OH group.
Huiqing Zhou et al., "m1A and m1G Potently Disrupt A-RNA Structure through the Intrinsic Instability of Hoogsteen Base Pairs" Nature Structural and Molecular Biology 23(9): 803–810.(2016)
لاحظ كيف أن الخصائص التى تستوفيها روابط هوجستين الناتجة عن ازالة ذرة الأكسجين متعلقة بعمليات أساسية كالتعرف على الحمض النووى و قراءته و نسخه و الحفاظ عليه و اصلاحه و هو ما يطعن بشدة فى امكان كون الرنا المفتقر لهذه الخصائص مخزنا أساسيا للمعلومات فى مرحلة سابقة من التطور من جهة و من جهة أخرى يرد على السؤال البوهيمى التطورى عن سر اشتراك كل الكائنات فى نفس الكمياء الحيوية لنقل المعلومات الجينية مفسرين ذلك بسلف مشترك أو مصمم مخادع...التفسير الواضح هو المصمم الذى اختار الكيمياء الحيوية المثالية للوظيفة. فى المقابل فان افتقار الرنا لهذه الخصائص ليس لكونه مرحلة سابقة من التطور لأنه عاجز أصلا عن العمل كمخزن طويل المدى للمعلومات بل لأن هذه الخصائص تعوقه عن أداء وظائفه كذاكرة مؤقتة أو كهيكل وظيفى عالى المرونة فهناك عمليات تعديل وظيفية methylation تقوم بمنع بعض الارتباطات الممكنة بين النيوكليوتيدات سواءا لأن الارتباط قد يؤثرعلى التطوى سلبا و يشوه الهيكل ثلاثى الأبعاد المطلوب أو لأن المطلوب أصلا هو منع التطوى و الهيكل ثلاثى الأبعاد حسب الوظيفة المطلوبة و مثل هذه التعديلات تقتضى أن لا يستطيع الرنا تجاوزها بروابط بديلة حتى لا تفشل – من جديد كل مكون مضبوط بدقة وفقا لوظيفته
As a result, whereas the DNA double helix can absorb damaged nucleotides (such as N1-methyldeoxyadenosine (m1dA) and N1-methyldeoxyguanosine (m1dG)) that are incapable of forming WC base pairs by forming HG base pairs, the same methyl marks, N1-methyladenosine (m1rA) and N1-methylguanosine (m1rG), acting as a post-transcriptional modification in RNA, block base-pairing altogether. These phenomena provide a direct mechanism for potently modulating the structure of the epitranscriptome. Our results indicate that HG-dependent DNA biochemical transactions may not be as readily supported in RNA duplexes, and they identify a unique dynamic property in B-DNA that may help enhance its ability to function as the repository of genetic information.
The greater instability of HG base pairs in A-RNA gives rise to a chemical switch in the form of m1rA and m1rG that can potently modulate RNA structure...For example, m1rA9 has been shown to stabilize the native structures of human mitochondrial tRNAs by blocking helical rA-rU WC base pairs that would otherwise stabilize alternative secondary structures (Fig. 6a). Likewise, m1rG37 next to the anticodon loop, which is highly conserved in most tRNAs that read the CNN codon[where N means any nucleotides], has been shown to prevent +1 frameshifting by blocking base-pairing between G37 and the first rC in the codon sequence (Fig. 6b). If RNA behaved similarly to DNA, such post-transcriptional modifications would simply create HG base pairs and fail to block base-pairing and to have their proper functional consequences.
The formation of stable mRNA secondary structure around start codons has been shown to reduce translational efficiency...it is possible that m1rA enhances translation in part by destabilizing secondary structure at the 5′ UTR near the start codons

الملفت فعلا هو أن نفس التعديل هو هو يعتبر تلفا يجب اصلاحه فى مخزن المعلومات طويل المدى بينما يعتبر ضرورة وظيفية فى الهيكل المؤقت و هو ما يشكل تناقضا واضحا فى امكان أحدهما القيام بالوظيفتين من قبل هيكل واحد كما فى فرضيات عالم الرنا
These modified bases block WC pairing because of steric collisions with the methyl group and because the methylation prevents one of the WC hydrogen bonds from forming (Fig. 3a). Both m1dA and m1dG occur in DNA after alkylation damage. In B-DNA, m1dA and m1dG are accommodated as m1dA-dT and m1dG-dC+ HG base pairs (Fig. 3a), which can in turn be recognized and repaired by damage-repair enzymes. m1rA and m1rG can also occur as a form of alkylation damage in RNA, but they are also highly conserved post-transcriptional modifications in tRNAs and rRNAs, and they play critical structural and functional roles, often by blocking WC base-pairing
و للمرء فعلا أن يتساءل ان كانت تعديلات الرنا بعد نسخه من الحمض النووى post-transcriptional هذه ضرورية للوظائف فى الرنا النقال و الرنا الرايبوسومى و هما أساسيان فى عملية فك الشفرة و ترجمة الحمض النووى الى بروتينات و يتم اعتبارهما من مخلفات عالم الرنا و أحد القرائن على وجوده فكيف كان عالم الرنا يعمل أصلا قبل "تطور" اليات تعديل الرنا و اضافة مجموعات الميثيل فى خلايا الرنا التى يزعمون وجودها كما سنرى فى الأسفل؟؟؟
أشارت الورقة السابقة أيضا الى أن أحد الفروق الجوهرية بين الدنا و الرنا قابلية الدنا الأعلى للانضغاط بفضل مرونته الناتجة عن ازالة مجموعة الهيدروكسيل و هذه المرونة الغير متحققة فى الرنا مهمة لعمليات الطى الفائق supercoiling المطلوبة لضغط مساحة تخزينه فى الكائنات الأكثر تعقيدا و التى تحتوى على حمض نووى أكثر ثم اعادة فتحه للوصول الى المعلومات فى داخله
In B-form DNA (B-DNA), the five-membered deoxyribose ring is flexible and favors the C2′-endo sugar pucker (Fig. 1a). In contrast, owing to the sugar 2′-OH group, the sugar in A-RNA is more rigid and adopts an alternative C3′-endo conformation (Fig. 1a). This conformation in turn brings the oxygen atoms (O5′ and O3′) adjoining sequential nucleotides closer, thus effectively compressing and rigidifying the A-form helix, widening its helical diameter, and displacing base pairs away from the helical axis. In addition, B-DNA and A-RNA differ considerably with respect to their deformability, and B-DNA is generally more flexible. The higher bendability of B-DNA than A-RNA is fundamentally important in many biochemical processes including the tight compaction of genome within the nucleus in higher-order organisms

المزيد من بصيرة الطبيعة و استعدادها لما هو قادم فى المستقبل من كائنات أكثر تعقيدا و لا يملك التطورى سوى قلب الاستدلال بالكلام الانشائى المعتاد بالقول بأن احلال الدنا محل الرنا هو الذى فتح الباب لتطور المخلوقات الأعقد فلا هو شرح كيف حدث الاحلال و لا كيف تطورت المخلوقات و لا حتى كيف استمرت حياة بالاعتماد على الرنا بعد كل ما رأيناه و سنراه من عدم ملاءمته و الأسوأ أننا سنقابل اعترافات منهم بأن دخول الدنا يمثل سم للرنا (ستمر علينا كوميديا سوداء عن فرضيات خلايا قديمة تعتمد على الرنا و بمناسبة الخلايا فحتى البكتيريا تحتاج الى الطى الفائق و بالتبعية مرونة الحمض النووى ناهيك عن وجود البروتينات التى تتعامل معه)
يظهر لنا من جديد مدى ملاءمة كل مكون لوظيفته من اللحظة الأولى و عدم امكان استبداله بشكل لا يسمح لا بانتخاب طبيعى من شئ اخر سابق و لا بمصادفة فى تفاعل كيميائى كان يمكن أن ينتج شيئا اخر و كعادة التطوريين ستجد دائما مزاعم عن امكان وضع مكونات أخرى من سكريات أخرى و لكن دائما ستظهر عيوب مقارنة بالمكون الأصلى كحدوث تأثيرات على مدى تقابل و ارتباط النيوكليوتيدات base pairing أو مقدار تراصها فوق بعضها stacking أو عدم القدرة على انشاء لولب مزدوج يوازن بين المرونة الحركية و الاستقرار فلا يطغى أحدهما على الاخر. ان شكل اللولب المزدوج المعتاد و الشهير يسمى B-dna هو فى الواقع أحد أشكاله الممكنة لكنه ليس الوحيد فتحت ظروف خلوية معينة كالجفاف مثلا أو عند وجود تسلسلات معينة (تتابعات من الحروف) أو أثناء عمل بعض البروتينات عليه يتعرض هذا الشكل لتغيرات هيكلية كأن ينضغط مثلا أو يتعرج أو يدور فى عكس الاتجاه المعهود A-Dna/Z-Dna و بالتالى يحتاج الى درجة من المرونة تسمح له بذلك ناهيك عن المرونة المطلوبة للف "السلم"ابتداءا لتحويله الى لولب مزدوج منغلق على نفسه للضغط و الحماية و اعادة فتحه عند الحاجة الى المعلومات داخله و فى المقابل فان زيادة المرونة بلا حساب قد تؤدى الى عدم تماسك اللولب المزدوج و ميوعة هيكله و عدم استقراره و الحاجة الى اليات أكثر للتعامل مع كثرة تنوعات بنيته بل و ربما ضعف قدرته على حماية النيوكليوتيدات داخله و الانغلاق عليها و تقريبها من بعضها بما فيه الكفاية لترتبط معا. بعبارة أخرى تحتاج الى موازنة دقيقة بين المرونة و الاستقرار للوصول الى المستوى المثالى optimization not maximization كما وصفته الدراسات و هو ما توفره حلقة الرايبوز الخماسية بيتا دى دون سواها لهذا اختارتها الطبيعة
optimization, not maximization, of base-pairing strength was a determinant of RNA's selection. Biological reasoning would emphasize that moderate base-pairing strength, as encountered in RNA and resulting from the high conformational flexibility of the ribofuranose backbone, was essential for the evolution of a rich diversity of nucleic acid-related biological functions. The observed base-pairing strength[much stronger Watson-Crick base pairing than DNA...consequence of the higher rigidity of pyranose as compared to that of furanose rings] in the family of the conformationally more rigid pentopyranosyl[six-membered pyranose ring] oligonucleotide systems provides a background of chemical facts in support of such an argument [all members of this family of RNA isomers were stronger Watson-Crick base-pairing systems than RNA itself]
Albert Eschenmoser “Chemical etiology of nucleic acid structure” Science Vol 284, Issue 5423 pp. 2118-2124 [Square brackets are quotations from elsewhere in the paper]
threose nucleic acid (TNA) is chemically simpler than RNA and has other properties that make it a plausible alternative to RNA. The lack of rotational freedom could have prevented them from becoming self-generating molecules...the functional groups attached to ribose are distantly located securing the free movement...whereas in chains of artificial nucleic acids (glycol nucleic acid [GNA]; threose nucleic acid [TNA]; peptide nucleic acid [PNA]), RNA functional groups would be in close proximity with glycol, threonine, and the peptide base, respectively
Gaspar Banfalvi "Ribose Selected as Precursor to Life" DNA AND CELL BIOLOGY Volume 39, Number 2, 2020 p. 177-186
Yonghang Run et al., "Formation and biological implications of Z-DNA" Trends in Genetics Review Volume 42, Issue 2 p163-176 February 2026
a much larger structural diversity of ‘six-membered’ versus ‘five-membered’ oligonucleotide duplexes. The potential for cross-talking between ‘six and six’ and between ‘six and five’ oligonucleotides further increases the available duplex structures...indicating the strong influence that modified nucleotides can have on duplex stability...Nature has selected the 1-β-d-ribofuranose nucleotides as building blocks for the genetic system, most probably based on their flexibility…Transitions between B- and A-forms of DNA through the influence of external agents such as proteins, water and counter-ions represent the common structural movement of a natural DNA duplex. This apparent structural continuum of natural nucleic acid conformations is reflected by the observation that it is often difficult to assign a DNA to a pure structural class when it is in complex with a protein.
Eveline Lescrinier rt al., “Difference in Conformational Diversity between Nucleic Acids with a Six-Membered ‘Sugar’ Unit and Natural ‘Furanose’ Nucleic Acids” Nucleic Acids Research Vol. 31, no.12 (June 15, 2003): 2975 – 89
و البحث الأخير بالذات لطيف جدا فبعد أن قارن تأثير اختلاف الحلقات على تراص الحروف/القواعد النيتروجينية المرتبطة بها فوق بعضها و قدرتها على القيام بحركات متعددة shift/slide/rise/tilt/roll/twist و تأثيره على سمات تقابل الحروف فى لولب displacement/inclination/tip/No. of nucleotides per turn خلص الى أن الطبيعة قد اختارت الحلقة التى تحد من المدى الحركى المتاح للولب مزدوج من الأحماض النووية بما يناسب استراتيجيات التحكم فى التعبير الجينى!!!

This conformational continuum is, only to a limited extent, accessible for interaction with natural nucleic acids (because of the five-memberedness of the furanose ring). It is clear that nature decided to work with only a small window of potential duplex structures, which is driven by thermodynamic considerations. Synthetic nucleic acids whose geometries fall beyond this accessible region are not suitable for control of gene expression (antisense and antigene strategies).
يعنى الطبيعة كانت تعلم أن التطور قادم فى الطريق و اللولب المزدوج قادم فى الطريق و الجينات قادمة فى الطريق و التحكم فى التعبير عنها و ضبطها قادم فى الطريق فاختارت المكونات الأمثل لذلك! أو أن المكونات المثالية لناسخ بدائى مزعوم (بفرض وجوده) تصادف أن تكون هى أيضا الأمثل لتعقيدات لا حصر لها لاحقة له و هذا طبعا من باب الصدفة و الحظ الحسن و ليس لأن هذه المكونات نحتت لهدف و غاية مستقبلية!
طبعا عندما تبحث فى تفسيرات التطوريين ستسمع جميع أنواع المبررات التى لا تفسر شيئا فمثلا هناك من سيذكر لك فوائد الحمض النووى فى حفظ المعلومات مقارنة بالرنا قائلا أن هذا هو السبب الذى انتقى من أجله الانتخاب الطبيعى الرايبوز منزوع الأكسجين دون أن يفسر كيف ظهر و دخل الى عمليات الخلية/الناسخ الذاتى بسيناريو معقول حتى يختاره الانتخاب (ستكون لنا وقفة فى الملحق مع بعض سيناريوهات المسخرة التى يطرحونها من خيالهم الشخصى و يسمونها علم) و هناك من سيتحدث عن ظهور الانزيمات التى تزيل الأكسجين من الرايبوز كمبرر دون أن يفسر كيف ظهرت أصلا التطويات البروتينية أو كيف استمرت الحياة مبنية على الرنا بسيناريو معقول – فقط لديك عصا التطور السحرية – قل أن كذا و كذا تطور أو انبثق أو ظهر فى تفاعلات الحساء البدائى أو ورث من سلف أو اذكر وظائفه ثم قل انتقاه من أجلها الانتخاب الطبيعى دون أن تشرح كيف نشأ ابتداءا أو قل جاء خطوة خطوة و لا يهم بعد ذلك عدم وجود انتخاب طبيعى أصلا قبل نشأة الناسخ الذاتى الأول من هذه المكونات المثالية و لا يهم أنه حتى فى وجود الانتخاب فان وظائف الشئ تبرر بقاءه و لكنها لا تفسر نشأته و لا يهم ان احتجت أن تفترض عالم ما قبل الرنا pre-RNA world لتبرر فرضية عالم الرنا ولا يهم كم الفرضيات و لا مدى معقولية السيناريو طالما تم وضعه فى ورقة علمية ما كان لها أن تمر فى أى منظومة محترمة لولا أنهم لا يجدون تبريرا اخر سوى البديل الأوقع الذى يكرهونه: التصميم! و عندما تضيف الى كل ذلك التكامل الوظيفى بين الرايبوز و الفوسفات و الذى يجعلهما الأفضل فى التفاعل لصناعة شكل الحلقة خماسية الأركان المثالى
They showed that although DAP[diamidophosphate] was able to phosphorylate all four sugars, it phosphorylated ribose at a much faster rate. Additionally, the reaction with ribose resulted exclusively in ring-shaped structures with five corners (e.g., 5-member rings), whereas the other sugars formed a combination of 5- and 6-member rings...And whereas the other three sugars got "stuck" at an intermediate point in the reaction, a large proportion of the ribose molecules were converted into a form that could likely react with a nuclear base to form a nucleotide.
"The sugar that sparked life: Why ribose was RNA’s first choice" ScienceDaily (July 23, 2025)
Harold A. Cruz & Ramanarayanan Krishnamurthy "Selection of Ribofuranose‐Isomer Among Pentoses by Phosphorylation with Diamidophosphate" Angewandte Chemie International Edition, Volume 64, Issue 35 (August 25, 2025)
تدرك أكثرو أكثر أن هذه المكونات ما كان لها أن تستبدل بأخرى و أنه من المستحيل تفسييرها بأحداث تاريخية عارضة historical contingency كان يمكن أن تنتج شيئا اخر بمكونات أخرى فقد نحتت خصيصا من أجل تلك الوظائف و ليكمل بعضها بعضا و هو ما سيظهر أكثر فى القسم التالى و نحن نناقش التكامل الوظيفى بين القواعد النيتروجينية
لماذا هذه القواعد النيتروجينية تحديدا؟
If you want to understand life, don’t think about vibrant, throbbing gels and oozes, think about information technology
Dawkins, R: 1991, The Blind Watchmaker, p. 112. (originally published by Longman Scientific & Technical, 1986.)
اذا أردت أن تفهم الحياة فكر فى تكنولوجيا المعلومات
-رتشارد دوكنز – كتاب صانع الساعات الأعمى
و الان نستعرض المكون الأخير و هى القواعد النيتروجينية و كالعادة نجد أن المسألة ليست مكونات دخلت التفاعلات فى فوضى الحساء البدائى و كان يمكن أن تطون أى شئ اخر و بالتالى لا يمكن تفسيرها الا بالسلف المشترك بل نجد أن النيوكليوتيدات المستخدمة منتقاة بعناية من بين النيوكليوتيدات الممكنة الأخرى لدرجة توقع بعض الباحثين أن هذه القواعد أو ما هو مماثل فى المواصفات يجب أن يكون موجودا فى أى حياة تنشأ فى الكون (ملحوظة: سيتم استخدام مصطلحات مثل حروف أو نيوكليوتيدات أثناء الشرح للاشارة الى القواعد النيتروجينية و هذا من باب العادة لا أكثر أما تقنيا فالقاعدة النيتروجينية جزء من النيوكليوتيدة/الحرف مثلها مثل الرايبوز و مجموعة الفوسفات)
Further, because of the unique abilities of purines and pyrimidines to interact with one another with particular specificity, these subunits, too, or something very similar to them, are likely to be common to life wherever it occurs
Pace, N.R. (2001) The universal nature of biochemistry. Proc Natl Acad Sci USA 98:805–808.
ستلاحظ معى فى استعراض هذه المواصفات شيئا ملفتا: البصيرة المستقبلية – و كأن "الطبيعة" كانت تعلم أنها ستستخدم هذه النيوكليوتيدات فى صناعة شفرة حمض نووى/دنا فى لولب مزدوج!!! (هذا على التسليم تنزلا بأن الحياة بدأت بشريط رنا منفرد ينسخ نفسه كما يزعمون و هو ما لا نسلم به أصلا)
تذكر كيف كتبنا فى المقدمة أن حروف الحمض النووى/الرنا تنقسم الى قسمين البيورينات A/G الأكبر حجما و التى تتألف من حلقتين bicyclic و البيورينات C/T/U الأصغر حجما و التى تتألف من حلقة واحدة momocyclic فوجود البيورينات و البيريميدينات بأحجامها المختلفة مهم جدا لصناعة اللولب المزدوج للحمض النووى - وجود حجمين مختلفين واحد صغير و الاخر كبير مهم حتى تكفى المساحة بين شريطى الحمض لاحتواء النيوكليوتيدات المتقابلة مع ضغط و اغلاق الشريطين لأن الارتباط يتم بطريقة صغير مع كبير و هذا التنظيم المكانى ليس مجرد حل هندسى أنيق لتخزين المعلومات و حفظها داخل هيكل مغلق تتوجه فيه حروف الشفرة نحو الداخل لتأمينها بل هو الية لضبط المعلومة المشفرة نفسها و حفظها من الأخطاء لأن ارتباط أحجام غير صحيحة معا (كبير مع كبير أو صغير مع صغير) يؤدى الى تشوهات و انبعاجات فى هيكل اللولب المزدوج يمكن انزيم النسخ polymerase (أو اليات البحث عن الأخطاء ان أفلتت من الناسخ) من التقاطها و تصحيحهاأو الى كون الحرفين بعيدين عن بعضهما و بالتالى غير قادرين على عمل الرابط الهيدروجينى hydrogen bond المطلوب. على صعيد اخر فان أزواج النيوكليوتيدات المتقابلة GC و AT/U ملائمة لبعضها البعض فى عدد الروابط الهيدورجينية بين كل زوج(3 و 2 بالترتيب) مما يجعل ارتباط بيورين و بيريميدين بشكل مختلف مثلا CA/GT أمر مشكل لهذه الروابط

و كالعادة يهرب التطوريون من ذلك بمحاولة طرح أسئلة لم نكتشفها بعد على غرار و لماذا هذه البيريميدينات و البيورينات تحديدا دون أخرى؟ لابد أن السبب هو أن الطبيعة لم تكتشفها أو أنها لم تدخل فى تفاعلات الحساء البدائى و كأن هذا الهروب يبرر نشأة الشفرة و اختيار نوعيات الحروف المناسبة ليأتى العلم من جديد باكتشاف السبب ليلتف التطوريون حول الاكتشاف و ينسبوه الى الانتخاب الطبيعى لتستمر طريقتهم الاستدلالية الثعبانية: لا نعرف لماذا اذن لا يوجد مصمم و التفسير هو السلف المشترك – نعرف لماذا اذن لا يوجد مصمم و التفسير هو الانتخاب الطبيعى ليظل التصميم مرفوضا من حيث المبدأ برغم أن الاكتشاف مرتبط بنظرية المعلومات!
أسلوب من أساليب تقييم اليات نقل المعلومات الرقمية و التشفير هو النظر الى المسافة hamming distance (∂) بين كل حرف/وحدة تشفير و الأخرى و المسافة هنا معناها عدد الأخطاء/التغيرات المطلوبة لتحويل وحدة من الشفرة الى أخرى فكلما زادت يزيد عدد الأخطاء المطلوبة لتحويل وحدة الى أخرى و تصبح الشفرة عصية على الخطأ و كلما قلت يمكن تحويل كلمة الى أخرى بسهولة و تصبح الشفرة رديئة الجودة معرضة لتبديل المعنى و تغير الكلمات بسهولة. لقد رأينا كيف أن حجم النيوكليوتيدة عنصر يساهم فى منع الأخطاء من نوعية ارتباط كبير مع كبير أو صغير مع صغير لكن هل يكفى بمفرده؟ و ماذا عن ارتباط كبير مع صغير اخر غير الذى يجب أن يرتبط به؟ اذن فمن منظور معلوماتى نحتاج الى الية رصد أخطاء اضافية لرفع كفاءة الشفرة. ان نمط الروابط الهيدروجينية بين الحروف مرسل/مستقبل Donor/Acceptor D/A يعمل بنفس طريقة التقاط الأخطاء فى المعلومات الرقمية و التى تترجم بيوكيميائيا الى مقدار التنافر بين النيوكليوتيدات الخاطئة فهناك حروف قد تتنافر معا فى موضع واحد من الروابط الهيدروجينية و أخرى فى موضعين و "الطبيعة"انتقت الحروف التى ستتنافر فى موضعين لأن التنافر الأقوى سيساعد أكثر فى تحديد الأخطاء و بالتالى فهذه الحروف ليست حادثة كيميائية بل ميزة تصميم – عفوا أقصد ميزة تطور
Brocklebank and Mac Dónaill : 2003, An ab initio Quantum Chemical Investigation of the Error-Coding Model of Nucleotide Alphabet Composition, Mol. Phys. 101:17.

U, C, G and aA (amino-adenine, an idealised form of A) form a subset of such a parity-code, belonging to the same (even) parity subset, where, intriguingly, the purine/pyrimidine size motif relates to the D/A pattern as a parity bit (Figure 4b). Given the centrality of parity in data transmission, the observation of a parity structure within the nucleotide alphabet is clearly of potential significance…These are rather striking results, and they strongly suggest that the selection of A, C, G and U/T result from evolutionary advantage, and do not represent a frozen accident
Donal A. Mac “Why Nature chose A,C,G and U/T: An error coding perspective of nucleotide Alphabet composition” Origins of life and evolution of the Biosphere 33: 433-55
error-coding theory...bringing the analytical power and formal descriptive facility of computer science to bear on this most fundamental of scientific issues…
In mixed parity systems the interpyrimidine or interpurine distances may be as little as one, and noncomplementary purine–pyrimidine associations may be opposed in just a single D/A position...and by the relatively weak repulsion between opposed lone-pairs...The energetics of nucleotide association therefore is such that a single mismatch is insufficient to ensure fidelity. By contrast, in a parity code the minimum distance between codewords is two. In a nucleotide context a minimum distance of two means that pyrimidines (and purines) differ from each other in the setting of two of the three D/A features, and attempted non-complementary pyrimidine–purine associations are opposed in two D/A positions...With two opposed positions the repulsion is considerably greater
The role of D/A patterns is therefore twofold, serving to bind associating complementary pairs, while simultaneously opposing non-complementary associations. Any set of complementary nucleotides is approximately equivalent with respect to the former, however, a parity code alphabet is optimal with respect to the latter, ensuring that the association of non-complementary pyrimidine–purine pairs will be opposed in two of three D/A positions… Error-coding analysis however suggests that mixed parity alphabets with interpurine or interpyrimidine distances of one have an inherently low fidelity.
Dónall A. Mac "A parity code interpretation of nucleotide alphabet composition" Chemical Communications Issue 18: 2062-63
لاحظ كيف يتكرر معنا دائما سيناريو كون المكونات ملائمة لهدف مستقبلى فمهما تحدث التطوريون عن الرنا المتطوى القادر على القيام ببعض الوظائف فان ارتباط الحروف ليتطوى الرنا شئ و كونها ملائما لعملية ضغط اللولب المزدوج فى مساحته الضئيلة و رصد أخطاء النسخ بتشفير معلوماتى شئ اخر تماما. ان كلامهم عن الانتخاب الذى اختار هذه الحروف فى عالم الرنا لأنها تساعد على دقة النسخ لا مبرر له سوى فرضية التطور ذاتها فالاحتياج الوظيفى متحقق فى الخلية دون أن تفترض شئ قديم أبسط لم يثبت وجوده خارج المعمل. أضف الى ذلك أن هذا التنافر غير كاف بمفرده بدون وجود شريط مقابل ليتشوه الهيكل و اليات تلتقط هذا التشوه و أخرى تصلح الخطأ (سنشرحها لاحقا فى المقال فى قسم: التكامل الوظيفى فى اللولب المزدوج) و كل هذا غير متوافر فى عالم الرنا المزعوم و كما سنرى لاحقا عند الحديث عن كارثة الأخطاء error catastrophe فان كائنات أكثر تطورا من عالم الرنا و تمتلك قدرات نسخ ذاتى أفضل و أدق و اليات تدقيق و مراجعة proofreading و اصلاح أخطاء لا يمكنها أن تصمد بالاعتماد على تنافر الحروف فقط ان تلفت هذه الاليات فتنافر الحروف هو الأساس الذى تعمل عليه هذه الاليات و لكنه ليس بديلا بدائيا عنها و هذا الأساس لا غنى عنه و لا يصلح شئ اخر كأساس بديل (و سنرى اثباتا اضافيا لذلك فى ملحق الرايبوزايم QT45 )
لقد قارنت الأبحاث بين نيوكليوتيدات أخرى ممكنة فى الحساء البدائى من حيث الحجم و مدى تكامل/تنافر مواضع الروابط الهيدروجينية و عدد النيوكليوتيدات فى الشفرة لتصل الى وجود أربع تصنيفات مختلفة للشفرات – نوع يزيد من الحروف/النيوكليوتيدات المستخدمة فيزيد السعة المعلوماتية و لكن تدخل فيه تنافرات أحادية كثيرة (مسافة صغيرة) فتقل دقته و يصبح عرضة للخطأ بشدة – نوع يفعل العكس فيقلل عدد الحروف و ينتقى الحروف التى بينها مسافات كبيرة و تنافرات شديدة فيقلل احتمال الخطأ و يزيد الدقة و لكن على حساب قلة الحروف و بالتالى قلة السعة المعلوماتية و هو ما وصفته الورقة بموازنة السعة المعلوماتية للشفرة مقابل دقتها tension between information density and replication fidelity - نوعان من الشفرات يوازنان بين الدقة و السعة المعلوماتية و لكن على حسب السمات الكيميائية للحروف المستخدمة فأحدهما Category II أعلى فى السعة المعلوماتية و أكثر مقاومة للأخطاء و من هذا النوع السعيد تأتى توليفة الحروف المستخدمة فى الشفرة الجينية لنصل فى النهاية الى نتيجة مفادها أن الطبيعة قد اختارت الحروف المثلى بطريقة تمنح الشفرة خاصية مقاومة الأخطاء بشكل بسيط لكن فعال simple but effective error-resistant properties و أنها قد استبقت نظرية أخطاء التشفير التى وضعها العلماء فى مجال المعلومات فى القرن العشرين (نعم هم يقولون هذا بالحرف فى الورقة ينسبون الذكاء الى الطبيعة و هو مذهب فلسفى يسمى panpsychism يتم تمريره كثيرا كجزء من تعريف العلم الطبيعى)
error-coding theory considerations inform us that not all combinations of patterns are equivalent with respect to error resistance. In a molecular context this suggests that not all combinations of nucleotides would be equally error-resistant, allowing selection pressure to select an optimal alphabet with respect to informatics...error-coding considerations show how a parity code structure might offer a replication fidelity advantage. The natural alphabet appears to be structured like a parity code, and it would appear that the error-coding theory proposed by Hamming in 1950 was actually anticipated by nature.
Dónall A. Mac "A parity code interpretation of nucleotide alphabet composition" Chemical Communications Issue 18: 2062-63


Check section: 3. Alphabet Categories: Information Content vs. Error-Resistance,
Specially 3.6 COMPARISON OF ALPHABETS
Donal A. Mac “Why Nature chose A,C,G and U/T: An error coding perspective of nucleotide Alphabet composition” Origins of life and evolution of the Biosphere 33: 433-55
نفس التطورى الذى يقول لك لا يوجد أثر للتصميم فى الكائنات الحية هو ذاته الذى يقول لك الطبيعة استبقت مجال التشفير و المعلومات لنظل ندور فى فلك خرافة صانع الساعات الأعمى علم و لكن المبصر ليس علم مهما أدى ذلك الى قصص خيالية اضافية كوجود شفرة سابقة مزعومة ليطورها الانتخاب الطبيعى أو قدرته على تطوير شفرة ابتداءا بفرض امكان نشأة واحدة من تفاعلات كيميائية لتبقى النتيجة الواضحة: المكونات المشتركة موجودة لأنها الأفضل للوظيفة و ليس لأنها موروثة من سلف مشترك
الناظر الى الجدول أعلاه ربما يجد شيئا غريبا: التصنيف الثالث يحمل مسافة/عدد أخطاء مطلوب لافساد التشفير أعلى من الثانى و مع ذلك ليس مقاوما للأخطاء error resistant كيف؟ لأن هناك نوعان من الأخطاء:
النوع الأول transitions و هذه التى تتبدل فيها النيوكليوتيدة بواحدة من نفس الحجم (بيورين يتبدل ببيورين اخر أو بيريميدين ببيريميدين) و بالتبعية لا يتأثر تقابل الأحجام صغير/كبير فى اللولب المزدوج و يكون الاعتماد فى كشف الخطا هنا على تنافر الروابط الهيدروجينية فقط
النوع الثانى من الأخطاء transversions هى تلك التى يتبدل فيها حرف بواحد من حجم مختلف (بيورين ببيريميدين أو العكس) و يكون جزء من الية كشف الخطا هو تشوه الهيكل بسبب ارتباط أحجام متشابهة. توليفة النيوكليوتيدات المستخدمة فى التصنيف الثالث تحتوى على نيوكليوتيدات ذات روابط هيدروجينية متكاملة مع نيوكليوتيدات لها نفس الحجم و بالتالى يمكن لحرفين من نفس الحجم أن يرتبطا معا بقوة تعوض أى انبعاج أو تشوه فى الهيكل ينتج عن ارتباط الأحجام الخاطئة. هذا يجعل هذا النوع من التوليفات من الحروف برغم دقته عرضة لنوع اخر من الأخطاء النابعة من حجم النيوكليوتيدات المرتبطة و ليس من تنافر الروابط الهيدروجينية.
CATEGORY III ALPHABETS...The increased resistance to associations between non-complementary purine/pyrimidine pairs comes at the expense of facilitating transversions, through perfectly complementary D/A patterns for associating purines or associating pyrimidines...Category II alphabets offer resistance to transitions ( ̄∂ = 2), through opposing association in two of three D/A positions; they also retain resistance to transversions ( ̄∂ = 2) by opposing such associations in one of three D/A positions in addition to the mechanical strain of transversions. These alphabets, having up to 8 letters, have a potential information density of 3 bits/letter. Category II alphabets are therefore optimal when information density and fidelity are considered together.
information in nucleotides is molecularly encoded in the hydrogen D/A patterns and purine/pyrimidine motifs. It is shown that in the natural alphabet the pyrimidine/purine motif is related to the D/A patterns as a parity bit. Nucleotide alphabets consisting of same-parity nucleotides have a replication fidelity
advantage so that the apparent inherent information structure which mimics a parity check code is unlikely to be a frozen accident.
اذن فالمسافة بين حرف/وحدة تشفير و أخرى تعتمد على مزيج من الحجم و تنافر/تكامل الروابط و كلا العاملين قد يؤثر على الاخر سلبا أو ايجابا. يظهر لنا واضحا هنا كيف تتكامل سمات متعددة ليس فقط على مستوى المكون الواحد بل على مستوى طريقة تفاعله مع باقى المكونات لتقود الى نتائج غير متوقعة بعيدا عن الطريقة شديدة السطحية التى يتعامل بها التطوريون – هناك مكونات أخرى ممكنة كان يمكن أن تكون أيها و بالتالى التشابه دليل سلف مشترك – لكن مهلا! لقد وصلنا الان الى 8 نيوكليوتيدات/حروف ملائمة لموازنة السعة المعلوماتية مع الدقة فلماذا انتقت "الطبيعة"4فقط مع أن 8 كان يمكن أن تقود الى سعة معلوماتية أكبر؟ هل هو من جديد السلف المشترك و الحوادث المجمدة فى الزمن؟ من جديد الاجابة: لا فقد نظرت الأبحاث فى حجم الشفرة لتجد 4 حروف هو الرقم المثالى
قارنت الأبحاث عدة خصائص ستتأثر بتغيير عدد النيوكليوتيدات سواءا فى مجال الابقاء على دقة النسخ و تقليل عدد الأخطاء مقابل قابلية الشفرة "للتطور" (التغير/التكيف مع التغيرات البيئية) أو فى مجال قيام الرنا بوظائفه و قدرته على التطوى الى شكل ثلاثى الأبعاد لتصل الى أن هذه الخصائص ستتأثر سلبا اذا زاد عدد الحروف و أن 4 هو العدد الأمثل لموازنة كل هذه المتطلبات
As with every enzyme, the enzymatic capacity of RNA rests on the three-dimensional positioning of functional groups...such as the cloverleaf structure of transfer RNA...three measures: the fraction of paired bases required to obtain the optimally folded structure, whether there are many or a few nearly optimal structures for the same sequence and the difference between the optimally folded structure and a completely unfolded one...In this test, the fittest alphabet was that with four bases...All three of these measures decrease with increasing alphabet size...expansion in size must be contained as it is probably offset by the increased risk of incorporating an incorrect base during replication.
As the alphabet size (N) increases, the accuracy of insertion of cognate bases decreases (which is affected by the difference in base-pairing energy between correct and incorrect pairs)...This by itself would select for a small alphabet size...there must be an optimal N (N*) that confers the highest fitness on ribo-organisms. Under a wide range of parameter values (such as temperature) N* = 4, as in the natural alphabet.
It might therefore be concluded that all theoretical investigations point in the same direction: a certain alphabet size (probably four) seems to be optimal as a compromise between stability and evolvability, between fidelity and catalytic efficiency, and between information density and error resistance.
Eörs Szathmáry "Why are there four letters in the genetic alphabet?" Nature Reviews Genetics Vol. 4 (December 2003)
طبعا الدوجما التطورية لا حدود لها و الهروب من الحاضر الى الماضى لا يتوقف...لدينا رنا فى الخلية الحية أمامنا يقوم بوظائف كثيرة و هذا بمفرده تفسير كاف لكن التطورى يجب أن يهرب الى أن هذه الوظائف نشأت فى عالم الرنا مستحيل الحدوث أصلا!لا حاجة لنا لافتراض عالم رنا لنفسر لأن الرنا الوظيفى موجود و فعال و مهم فى الخلية بعيدا عن خيالات كونه بقايا من منظومة قديمة تفترض أن قوى التطور الخارقة التى أبدلت كل منظومة الرنا من تشفير بالحمض النووى و تحفيز تفاعلات و أعمال وظيفية أخرى بالبروتينات عجزت عن تبديل هذه البقايا!!
اذن فالعدد الأمثل لحروف الشفرة 4 و عدد الحروف المرشحة لكونها المثلى لموازنة السعة المعلوماتية مع قابلية الخطأ 8فلماذا هذه الأربعة تحديدا؟ حادثة مجمدة؟ من جديد لا فالأبحاث قارنت النيوكليوتيدات الثمانية من حيث صفات متعددة أخرى لتجد أن الحروف الأخرى لا تستوفى هذه الخصائص
chemical stability (the base should not readily decompose); thermodynamic stability (new base pairs should not destabilize nucleic-acid structures); enzymatic processability (polymerases should accept the base pairs as substrates, catalyse addition to the primer and be able to carry on the process); and kinetic selectivity (ORTHOGONALITY to other base pairs)...a new base pair that satisfies all four conditions (chemical and thermodynamic stability, enzymatic processability and kinetic selectivity) has not yet been achieved
طبعا لعب الخيال التطورى دوره فى محاولة الهروب من هذه الحقائق كالزعم مثلا بأن عدم توافق بعض النيوكليوتيدات مع انزيمات الخلية سببه أن التطور أنتج هذه الانزيمات خصيصا من أجل نيوكليوتيدات الشفرة الحالية دون أى اثبات لأن التطور أصلا يمكنه أن ينتج اليات الخلية الحالية و هى عادة التطوريين فى دعم قصصهم الخيالية بقصص خيالية أخرى لصناعة سيناريو يبدو محبوكا و متسقا دون دليل على حدوثه فى الواقع لكن حتى مع هذا بقيت سمات أخرى فى النيوكليوتيدات الغير مستخدمة توضح عدم مناسبتها للوظيفة مثل عدم استقرار هذه النيوكليوتيدات على هيكل محدد و قابليتها لاتخاذ أشكال مختلفة غير ملائمة tautomeric instability أو سهولة تحللها hydrolysis/aminolysis أو مقدار الطاقة فى معالجتها أو مقدار توافقها مع العمليات التى تخضع لها النيوكليوتيدات كالفسفرة ناهيك عن كفاءة المنتج الصادر عنها فى بيئة الخلية الحقيقية أو توافقها مع باقى النيوكليوتيدات المستخدمة (تذكر كيف فى حالة الأحماض الأمينية كان أحد العوامل الهامة هو تكامل الحزمة مع بعضها البعض و مع المسارات البيوكيميائية و عمليات الانتاج وتدوير المكونات الموجودة فى الخلية بالفعل لأن المقارنة تكون بين فائدة الحرف الاضافى/البديل مقابل تكلفة انتاجه) ناهيك عن أننا وسط كل هذا نتحدث دون أن نثبت ميزة حقيقية لحرف مختلف أو اضافى بل حتى الان عيوب
Check section: 4. Chemical and Coding Theory Constraints in the Even Parity Alphabet
And section: 5. The Odd-Parity Alphabet
Donal A. Mac “Why Nature chose A,C,G and U/T: An error coding perspective of nucleotide Alphabet composition” Origins of life and evolution of the Biosphere 33: 433-55
assess the template and enzymatic capabilities of macromolecules that are built from bases of the extended alphabet...The efficient use of a new base pair —when one is ultimately found that meets all criteria — will require not only efficient polymerase action but also enzymatic phosphorylation of the nucleoside analogues
Eörs Szathmáry "Why are there four letters in the genetic alphabet?" Nature Reviews Genetics Vol. 4 (December 2003)
potential biosynthetic relationship or various other factors...biosynthetic pathways and that “network closure”" the notion that all processes inside a cell should be linked and share common resources for efficient coordination of metabolism, may have been important in the adaptive evolutionary construction of biochemistry
Melissa Ilardo et al., "Adaptive properties of the Genetically encoded Amino Acid Alphabet Are inherited from its Subsets" Scientific Reports volume 9, Article number: 12468 (2019)
لكن ماذا عن الحرف المختلف بين الدنا و الرنا. يستخدم الرنا اليوراسيل مكان الثايمين و الحرف البديل هو بيريميدين (راجع المقدمة) ينتمى الى نفس فئة حجم و نفس نمط الروابط الهيدروجينية للحرف المستبدل حتى يحل محله بكفاءة و لكن لماذا الاستبدال أصلا؟ ذهبت احدى الدراسات الى أنه طالما أن اليات قراءة و نسخ الشفرة تعتمد على تمييز السمات الفيزيائية و الكيميائية للحروف فان هذه الحروف يجب أن تتشابه الى درجة ما حتى تتعامل معها نفس الاليات المشتركة و لكن فى نفس الوقت تختلف بما فيه الكفاية حتى لا يختلط الأمر على هذه الاليات فى قراءتها و تمييزها عن بعضها البعض و لا يكون هناك ميل خاص للخلط بين حروف معينة (نفس فكرة مسافة الخطأ hamming distance التى ناقشناها بالأعلى و لكن هذه المرة حتى على شريط واحد بدون وجود لولب مزدوج و نفس فكرة موازنة الكثير من المتطلبات معا)

In the genetic code all sequence recognition mechanisms are based on physicochemical interactions between different polymerases, co-factors and the macromolecules DNA and mRNA. Hence similarities and differences relevant to the genetic coding should in some way be reflected by the physicochemical properties of the bases comprising the code. On one hand the bases must share some basic properties in common to work with in the same reading mechanism. On the other hand they must differ sufficiently in other respects such that the code can be unambiguously read. Properties of the five bases which are known to be important in DNA and mRNA are their stability, their ability to form hydrogen bonds and their tendency to form specific base pairs of similar sizes… the elements of an ideal genetic code should be (i) as dissimilar as possible within a common reading mechanism and (ii) equally dissimilar to each other.
Jorgen Jonsson et al., “The evolutionary transition from uracil to thymine balances the genetic code” Journal of Chemometrics 10, no.2(1996): 163
قامت الدراسة هنا بتقييم السمات الرئيسية principal properties (Pps) للحروف بطريقة شبيهة بدراسات الأحماض الأمينية التى ذكرناها فى مقالنا السابقhydrophobicity, polarity, polarizability ,size (steric bulk), hydrogen bonding فوجدت أن حروف الحمض النووى الأربعة كلهم يمثلون أقصى درجة تباعد فى السمات بينهم دون الخروج عن الاطار الذى يسمح بالتعامل معهم بنفس الاليات و هذا هو الوضع المثالى لأى شفرة جينية لتسهيل و تسريع قراءتها و تقليل الأخطاء
The use of four different bases that systematically and symmetrically differ from each other in three important and independent physicochemical aspects is probably a very efficient way to obtain a stable code. The fact that the smallest possible number of properties is used to give each base a unique identity probably also optimizes the efficiency and speed with which a particular base may be recognized… the fidelity of DNA replication will be enhanced
جميل جدا – لازلنا نواصل اثبات مثالية المكونات حتى على مستوى شريط منفرد و لكن لازلنا لم نفهم بعد ماذا عن يوراسيل الرنا؟ وجدت الدراسة أن هناك تشابه عالى نسبيا بين اليوراسيل و السايتوزين و بالتبعية فان حذف اليوراسيل من الشفرة الدائمة يباعد بين مكوناتها hamming distance ويحسن دقتها بنسبة 50% بينما على مستوى الرنا كمخزن معلومات مؤقت لا تحتاج الشفرة الى اهدار موارد لوضع مجموعة ميثيل اضافية على اليوراسيل لتحويله الى ثايمين
methylation of U to yield T (5 -methyluracil).The biological consequences of these changes are, however, considerable… The principal properties of the bases in DNA are seen to be distributed in a close-to-perfect tetrahedral shape. The corresponding scores for the bases of mRNA are also distributed in a tetrahedral shape, but the two pyrimidines C and U are considerably closer to each other…By substituting U for T, the variance of the score distances is reduced by almost 50%.The tetrahedron becomes more regular and as a consequence the chemical integrities of all four bases are simultaneously strengthened… Thus greater effort is spent in maintaining the integrity of the master copy than of its working copy. This appears to be an appropriate compromise, considering the product of energy and time required to synthesize and maintain these codes.
لاحظ أنهم يتعاملون بطريقة السحر التطورية اياها: ذكر فوائد الشئ كمبرر لظهوره – لا بأس فهو يعنى المزيد و المزيد من المثالية و ملاءمة كل شئ لوظيفته و المزيد من الطعن فى قدرة الرنا على العمل كمخزن معلومات طويل الأمد لكن لازالت هناك نقطة لطيفة فى الدراسة السابقة. فى بداية الدراسة ذكروا أنهم يبحثون عن تفسير اخر غير التفسير السائد لتبديل الحرفين فما هو التفسير السائد و لماذا بحثوا عن بديل له؟ ان السايتوزين يتحلل الى يوراسيل و هناك انزيمات متخصصة لازالة هذا الخطأ فلو استمر اليوراسيل كحرف فى الشفرة لقامت هذه الانزيمات بازالة اليوراسيل الأصلى (بديل الثايمين) مع اليوراسيل الناتج عن السايتوزين المتحلل ولانتهى الأمر عمليا بشفرة من ثلاث حروف فقط سايتوزين و جوانين و أدينين. المشكلة أن نظرية التطور تقوم دائما بافتراض (وليس اثبات) عدم وجود تصميم أو بصيرة أو هدف و افتراض (و ليس اثبات) التدرج و بالتالى التطور لا يملك بصيرة ليعرف أن اليات الاصلاح هذه ستظهر لاحقا ليعدل حروف الشفرة ليمكنها العمل و هذا هو سبب رفضهم لهذا التفسير برغم اتساقه التام مع البيانات – لأنه على حد تعبيرهم teleological ينم عن الغاية و البصيرة و الهدف.
The switch from U to T has also been explained in terms of chemical instability. This is based on the observation that C can undergo spontaneous deamination at a low but significant rate and thereby convert to U. Thus the U-to-T switch has been suggested to be advantageous since it allowed the development of the enzyme uracil-DNA glycosylase (UNG).This enzyme excises all Us (i.e. degraded Cs) found in DNA, subsequently allowing other components in the DNA repair machinery to act, thereby preventing the genome from a potentially harmful enrichment of false Us. This repair system could obviously not function if uracils were a normal component of DNA. This teleological explanation of the U-to-T switch, however, implies that evolution could foresee the future possibility of developing such a repair system. We present a novel, alternative rationale for this evolutionary transition, namely that of balancing the genetic code in a property space
هذه المنطلقات – رفض الغائية و الهدف و التصميم – هى الأساس الفلسفى الذى يفسرون على أساسه كل البيانات العلمية و لكن عادة لا ينطقونها بهذه الصراحة لذا تذكر مثل هذه الأمثلة كلما قال لك أحدهم لا شئ فى البيولوجيا يدل على التصميم و الغاية – الحقيقة أن كل شئ يشير الى ذلك و لكنهم يتأولون البيانات بطرق ملتوية و اذا أزلت العقيدة الفلسفية الباطلة القائمة على افتراض عدم وجود تصميم من رأسك لوجدت أن كل هذا دليل اضافى على التصميم و البصيرة و ملاءمة المكونات لأهدافها فالرنا لا يحتاج الى كلفة أعلى و لا يشكل له تحلل السايتوزين أى مشكلة أصلا لأنه مخزن قصير الأجل بطبيعته بينما الثايمين الأعلى كلفة فى الانتاج سماته ملائمة لمخزن المعلومات طويل الأجل الدنا و الية الاصلاح التى تعمل عليه (التى اخترعها التطور طبعا)

صورة للمسارات البيوكيميائية التى اخترعها التطور حتى يستقل و يعتمد على نفسه فى انتاج المكونات بعد أن دخلت الى الحياة من تفاعلات كيميائية فى بيئة الأرض البدائية – اخترع الانزيمات و العوامل المساعدة التى ستصنع الثايمين باضافة مجموعة ميثيل الى اليوراسيل طبعا بعد أن اختراع مسارا بيوكيميائيا لصناعة اليوراسيل ذاته (غير موضح فى الصورة) – لاحظ كيف تتقاطع المسارات البيوكيميائية للمكونات المختلفة سواءا نبيوكليوتيدات أو أحماض أمينية و تتكامل. الدليل على أن التطور "اخترع"كل هذا؟ سهلة اذكر فوائد المنتجات النهائية
Cytosine deamination results in mutagenic U:G mismatches that must be excised. The repair system, however, also excises U from U:A “normal” pairs. It is therefore crucial to limit thymine-replacing uracils...De novo biosynthesis of thymine is an intricate and energetically expensive process that requires dUMP as the starting material and a complex array of two enzymes and cofactors (Figure 1B). It is therefore straightforward to ask: is there any specific reason that justifies this costly and seemingly equivalent replacement of uracil by thymine in DNA?...For short-term storage of genetic information, as in modern RNAs, cytosine deamination rates do not pose a serious problem; therefore uracil does not need to be discriminated against.
Beata Vertessy and Judith Toth “Keeping Uracil out of DNA: Physiological role, Structure and Catalytic Mechanism of dUTPases” Accounts on Chemical Research 42, no.1 (2009): 97 – 106
لاحظ أن عملية التحويل من يوراسيل الى ثايمين تبدو فى الصورة ملخصة كخطوة واحدة و لكن الية عمل انزيم التحويل و خطوات تدوير و انتاج عوامله المساعدة أعقد بكثير من اختزالها فى "سهم" (انظر الصورة أدناه لخطوات التفاعل)

مسار انتاج اليوراسيل بعد أن كان الناسخ الذاتى الأول يحصل عليه من البيئة و الذى يجب وجوده قبل تحويله الى ثايمين

Donald Voet, Judith G. Voet and Charlotte W. Pratt "Fundamentals of Biochemistry: Life at the Molecular Level" 5th Edition (2016) p. 809, 819
لاحظ من فضلك أن كل الكلام السابق عن ملاءمة الرنا لوظيفة المخزن المؤقت فقط و تشابه السايتوزين مع اليوراسيل و تحلل الأول الى الثانى لها تبعات كبيرة على فرضيات عالم الرنا و الناسخ الذاتى الأول المزعوم و امكان نشأة هذه المنظومات بما فيها من تكامل وظيفى تدريجيا و سنتعرض لهذا أكثر فى الملحق
و على ذكر تكامل المسارات البيوكيميائية و تقاطعها معا فالقواعد النيتروجينية لا تستخدم فقط فى التشفير بل أيضا كمكون لوسيلة لنقل الطاقة مثل ATPأدينوسين ثلاثى الفوسفات و قد حاولت ورقة متعلقة بأصل الحياة دراسة المصدر "التطورى"للأدينوسين ثلاثى الفوسفات فاذا بها تقر بأن مصدر الطاقة الملائم يجب أن يكون مركب من أقارب نيوكليوتيدات (حروف) الحمض النووى لأسباب كيميائية متعلقة بصناعة شريط الحمض
A partial explanation is that ATP links energy metabolism with genetic information. ATP-coupled monomer activation promotes the polymerisation of macromolecules, including RNA, DNA, and proteins. Unlike the simple phosphorylation of intermediary metabolites, the leaving group during nucleotide polymerization is pyrophosphate (PPi)...The hydrolysis of PPi renders these steps strongly exothermic, if not practically irreversible—a ratchet towards polymerization. Only nucleoside triphosphates can release PPi while still retaining a phosphate for the sugar-phosphate backbone of RNA and DNA, or for amino acid activation.
Silvana Pinna et al., “A Prebiotic Basis for ATP as the Universal Energy Currency, PLoS Biology 20 (October 4, 2022): e3001437
يضيق هذا خانة المركبات الكيميائية الملائمة بشكل ضخم بعيدا عن خرافات "كان يمكن أن يكون أى مركب اخر"ثم حاولت الورقة تحديد لماذا الأدينوسين بالذات و دخلت كالعادة فى خرافات توافر المكونات المؤدية اليه فى بيئة الأرض القديمة و لكن بعيدا عن هذه التبريرات الواهية فالورقة اعترفت باختلافات تفاعلاته الكيميائية عن الحروف الأخرى unusual chemistry in water و بفشل بعض التفاعلات التى تمت تجربتها على الحروف الأخرى و الدلالة الكبيرة لانتشار مشتقات الأدينين فى عمليات الأيض الأخرى لدرجة أن قالت بوضوح هذه لا يمكن أن تكون صدفة من صدف الحساء البدائى و لا حتى انتخاب طبيعى
In our view, the prominence of ATP is unlikely to reflect a frozen accident, as adenine nucleotide cofactors are also ubiquitous across intermediary metabolism, including the ancient cofactors NADH, FADH, and coenzyme A...This implies that ATP could have become the universal energy currency of life not as the endpoint of genetic selection or as a frozen accident, but for fundamental chemical reasons, and arguably in a monomer world before the polymerization of RNA, DNA, and proteins.
طبعا الورقة حاولت وضع الأمر فى سياق الحتمية الكيميائية و هو كما وضحنا اعتراف باختلاف بمثالية الخصائص الكيميائية للأدينين لذا فالنتيجة ببساطة هى اختيار المصمم للمركب المناسب للوظيفة. النقطة الأخرى هى أن التطورى عادة ينتقل بسلاسة منقطعة النظير من "كان فى الحساء البدائى"الى "اخترع التطور الطريقة البيوكيميائية لانتاجه" و لتصور مدى لاواقعية هذا الكلام الانشائى انظر الى المسار البيوكيميائى المطلوب لتصنيع أول البيورينات. – كل خظوة بانزيم مستقل (مع ملاحظة أن الجملة عديمة المعنى التى يلقيها التطوريون فى هذه المواقف: كل انزيم كان يفعل شيئا اخر ثم تم تجنيده/استقطابه الى هذا المسار جملة لا تفسر شيئا بفرض صحتها أصلا – تعدد وظائف الشئ لا يشرح كيف نشأ و "انتقاه الانتخاب الطبيعى" جملة لا تشرح كيف نشأ لينتقيه أما فرضيات أن مصيدة الفئران كانت مثقل ورق ثم مثبت ربطة عنق فقائلها قبل سامعها يعلمون أن مصيدة الفئران لم تنشأ بهذه الطريقة و أن القدرة على اختراع قصة تبدو ظاهريا متناسقة لا تلغى تلك الحقيقة)

هذا المسار هو الذى يصل بالخلية الى مرحلة انتاج Inosine monophosphate (IMP) و نقطة تفرع انتاج البيورينات - بعدها يتفرع المسار الى فرعين فرع يحول imp الى جوانوسين G و اخر الى أدينوسين A و لاجقا تتم فسفرة المنتجات

طبعا العملية كلها منظمة وفقا لاحتياجات الخلية بعيدا عن فكرة مجرد توافر المكونات فى محلول خاصة و أنها تقاطع مع انتاج عملات توليد الطاقة

Donald Voet, Judith G. Voet and Charlotte W. Pratt "Fundamentals of Biochemistry: Life at the Molecular Level" 5th Edition (2016) p. 804, 806, 807
و من لطائف الكتاب السابق أنه يطرح أسئلة على الدارسين من طراز لماذا يبدو عقلانيا/منطقيا make sense أن تعمل المنظومة بهذه الطريقة ثم ينسب هذه العقلانية الى التطور و الطبيعة!
و من الصفات الملفتة جدا التى توضح ملاءمة هذه النيوكليوتيدات تحديدا دون غيرها و حاجة الطبيعة الى بصيرة مستقبلية حتى تعتمد عليها هو طريقة تفاعلها مع الأشعة فوق البنفسجية فمن جهة لها خصائص مقاومة لتأثير الأشعة الضار و من جهة أخرى الطول الموجى الأكثر عرضة لأن يؤذيها هو بالمصادفة الطول الموجى الذى يصده الأوزون!
nature has developed a variety of adaptations to control and limit the photochemical damage that might otherwise be initiated by the energy-rich excited states formed by the absorption of UV photons. Enzymatic repair, for example, actively reverses certain kinds of photochemical damage. Equally important safeguards are associated with the intrinsic photophysical properties of the bases themselves. The lowest energy transitions of the bases are located in a spectral region where the UV solar flux is significantly attenuated by the Hartley absorption continuum of ozone. Further protection comes from remarkably efficient relaxation pathways for electronic energy...These remarkable photophysical properties may be a further reason nature entrusted the genetic code to these compounds
Jean-Marc L. Pecourt, Jorge Peon, and Bern Kohler "Ultrafast Internal Conversion of Electronically Excited RNA and DNA Nucleosides in Water" Journal of the American Chemical Society Vol 122/Issue 38 (2000) p. 9348
من جديد نموذج للأمور التى لن ينفع فيها انتخاب طبيعى فى عالم ما قبل الرنا و لا عالم ما قبل قبل الرنا لأن عامل االانتقاء المفترض (الأوزون) لن يظهر الا فى المستقبل بعد ظهور الخلايا و التمثيل الضوئى الذى ينتج الأكسجين ما لم يكن الانتخاب الطبيعى يعرف المستقبل و أن طبقة الأوزون قادمة فينتقى الحروف التى يحميها الأوزون ثم يحافظ عليها بشكل ما حتى يظهر الأوزون. اختتمت الورقة بأن تقول أن هذه الخواص جعلت الطبيعة تأتمن هذه المركبات الكيميائية على الشفرة الجينية و طبعا لن نتعجب ان عارض التطورى كل قصصه السابقة ليخترع قصة لنشأة الأكسجين مبكرا أو يبتكر الية جديدة يضيفها الى فيلم الحساء البدائى الخيالى حتى ينجح "معمل الطبيعة"فى حماية النيوكليوتيدات من الطول الموجى المحدد الذى سيحمى منه الأوزون لاحقا و هذا ليس غريبا عليهم فهم قوم يعتبرون قدرتهم على تأليف سيناريو ما دليل على حدوثه.
النتيجة الان أن الحروف المستخدمة تمثل حالة مواءمة بين متطلبات كثيرة pareto optimality و هو مفهوم مهم فى عمليات التصميم التى يقوم بها البشر يشير الى الحالة التى لا يمكنك فيها تحسين أى جانب دون التأثير سلبا على جانب اخر
"Multi-Objective Optimality in Biology" in Uri Alon "An introduction to systems Biology, Second Edition" (2019)
ما بين تشفير معلومات و تقليل أخطاء و تكلفة انتاج و ملاءمة عمليات بيوكيميائية كثيرة و القدرة على اتخاذ هياكل وظيفية ثلاثية الأبعاد و الاستقرار الكيميائى و مقاومة التلف لذا فان الاعتراضات من طراز "هناك منظومات أخرى ممكنة أعلى فى تشفير المعلومات"أو "المنظومة لا تمنع أخطاء النسخ بنسبة 100% اذن هى غير مصممة"هى كلام لا يقول به عاقل غير مؤدلج بل و يتجاهل أن منع التغيير تماما معناه الحكم على الكائنات الحية بالفناء لعدم قدرتها على التكيف و التنوع لمواجهة تغيرات البيئة
و فى النهاية لا يتبقى أمام التطورى الا تشغيبات على غرار "لماذا بعض الخطوات البيوكيميائية قد تستخدم حرفا بديلا فى عملة الطاقة"أو "و لماذا توجد فيروسات قد تستخدم حرفا بديلا فى الشفرة؟"و كأن هذا ينفى كل الاكتشافات السابقة و كأن هذا دليل على نظريته و هو ذاته عندما نكتشف لماذا سيعود لينسبها لنفس النظرية التى يدللون عليها بالشئ و عكسه! بل ان التطورى مهما حاول أن ينسب هذه الظواهر الى خيالاته الشخصية ككونها بقايا من فوضى كيمياء الأرض الأولى أو من شفرات سابقة أو مسارات بيوكيميائية قديمة سيصطدم بالواقع القائل بأن هذه الحروف تتطلب انزيمات متخصصة للتعامل معها فالحرف الفيروسى البديل Z مثلا هو عبارة عن أدينين معدل 2-aminoadenine يحتاج انزيمات خاصة لتعديل الأدينين الأصلى و بلمرته polymerase لادخاله فى الشفرة أثناء نسخ الحمض النووى و تدمير الأدينين الحر حتى لا يدخل مكانه! بل انه و بالنظر الى ما سبق من اكتشافات و استقراء نتائجها فعلى الأرجح سنكتشف كالعادة أن هذا البديل موجود فى هذه الحالة لهدف و وظيفة متخصصة لنعود من جديد الى مربع "لماذا اختارت الطبيعة كذا و كذا؟"و الذى يعنى ببساطة اما ظهور و استخدام المكونات المثلى من اللحظة الأولى أو اضافة المزيد من القصص الخيالية عديمة الأدلة عن عوالم سابقة و كيمياء حيوية سابقة غير موجودة اليوم ظهرت فى الماضى ثم تم تعديلها. هكذا تطلق هذه العبارات بمنتهى البساطة مع أن هؤلاء العلماء أنفسهم يمضون شهورا أو حتى أعواما بعلومهم و خبراتهم و أجهزتهم لتعديل منظومة التشفير أو اضافة حروف/نيوكليوتيدات أو أحماض أمينية اليها ليخرجوا عليك بعدها مستدلين بما فعلوه بالتصميم على أفعال أخطاء النسخ و التفاعلات الكيميائية
ان أسهل ما يحفظه التطورى هنا هو الديباجات الانشائية المعتادة عن التدرجية كأن يقول أن فكرة تصحيح الأخطاء لم تكن ضرورية فى بداية الحياة مع الناسخ الذاتى الأول و أنها تطورت تدريجيا بعد ذلك بل و ربما يزيد الطين بلة بالتظاهر بأن كثرة الأخطاء فى البداية كانت دافعا للتطور – سنستعرض خطأ هذا الكلام عند الحديث عن كارثة الأخطاء بالأسفل و لكن تذكر الان أن أى تطور تدريجى لاحق يعتمد بشكل كلى على وجود النيوكليوتيدات الصحيحة الملائمة لعملية كشف الأخطاء من البداية من حيث نوعها و عددها فهل كان التطور يعرف أنه فى المستقبل سيطور عملية تصحيح الأخطاء ليخبر الطبيعة أن تصنع الناسخ الأول من المكونات المطلوبة نوعا و عددا؟ هل كان يعلم أنه فى المستقبل سيطور مسارات بيوكيميائية لانتاج الحروف و لغيرها من الوظائف حتى ينتقى الحروف التى تتناسب مع عمليات الخلية الأخرى؟ (انظر مثلا حديثنا عن ATP و استخدام قاعدة نيتروجينية من جهة و مجموعات فوسفور من جهة كمصدر مثالى للطاقة) بل ان عملية نسخ لولب مزدوج باستخدام كل شريط كقالب لصناعة شريط جديد مقابل تعتمد بداهة على ارتباط النيوكليوتيدات المتقابلة مع بعضها البعض بدرجة عالية من الدقة بأقل قدر ممكن من الأخطاء أو من عدم الاستقرار فحتى من أراد تجاهل تصحيح الأخطاء و حتى من أراد افتراض بدء النسخ الذاتى بشريط منفرد سيجد أن اختيار المكونات من البداية مبنى على مثاليتها لعملية نسخ لولب مزدوج قادمة فى المستقبل – من جديد و كأن التطور كان يعرف أن الشريط المنفرد سيتحول الى شريطين مرتبطين فى المستقبل!
سيحكى التطورى قصصا طويلة من رأسه و بلا دليل عن وجود نواسخ ذاتية كثيرة بمكونات مختلفة و هذا الناسخ هو الذى تصادف أن ربح سباق البقاء و بالصدفة البحتة اتضح أن سماته ملائمة لوظائف و هياكل و عمليات بيوكيميائية لن تظهر الا فى المستقبل و ان سلمنا جدلا بهذا مع أنه وفقا لكل ما نرى غير ممكن فما هذه الصدفة العجيبة التى جعلت مكونات محددة توازن كل هذه الصفات و المتطلبات المستقبلية معا و كأنها نحتت خصيصا لتكون أحجار بناء الحياة و ما لزوم تفسير السلف المشترك الان و الكلام عن عدم وجود أى مبرر لمصمم لينتقى هذه المكونات الا ليخدعنا و يوحى لنا بالتطور؟ ان طبيعة المكونات المثالية و ملاءمتها لأمور مستقبلية أعقد بكثير من مجرد النسخ الذاتى اضافة لطبيعة الرنا سواءا من حيث الرايبوز أو اليوراسيل التى رأينا كيف تجعله عاجزا عن ما ينسب اليه كلها أمور فعلا تصرخ فى وجوهنا قائلة أنه فعلا هناك من يحاول أن يخدعنا ليوحى لنا بالتطور و لكنه ليس المصمم!
-انتهى الجزء الأول من المقالة و الخاص باحاد المكونات و يتبعه الجزء الثانى ان شاء الله و الخاص بنتائج اجتماعها (اذا مر عليك شئ مكتوب أننا سنناقشه فى قسم لاحق من هذا المقال و لم تجده فهو فى القسم الثانى ان شاء الله)


Comments