أيقونات التطور 2: شجرة الحياة
- Illidan Stormrage

- Jan 10, 2021
- 70 min read
Updated: May 21, 2025
The relationships between major groups within an emergent new class of biological entities are hard to decipher and do not seem to fit the tree pattern that, following Darwin’s original proposal, remains the dominant description of biological evolution
Eugene V. Koonin, “The Biological Big Bang Model for the Major Transitions in Evolution,” Biology Direct 2 (2007): 1–17,
العلاقات بين المجموعات االرئيسية من الكائنات الحية صعبة الفهم و لا يظهر أنها تتفق مع شكل الشجرة المتفرعة الذى تتوقعه نظرية التطور
-من مراجعة علمية لنموذج ظهور الكائنات بين الواقع و نظرية التطور
To suggest that this "data" can be handed over to non-specialists so they can make up their own minds is to profoundly misunderstand the nature of science.
karl Giberson "On the Integrity of Science: A Response to Bill Dembski" p.5
ان اقتراح أن يتم عرض البيانات على الناس ليقرروا بأنفسهم يعتبر جهل بطبيعة العلم
كارل جايبرسون مناظر تطورى و كاتب كتب لاقناع الناس بالتطور
هذه الجملة العجيبة قالها ردا على عالم الرياضيات ويليام دمبسكى بعد أن انتقد كتابه المشترك مع التطورى فرانسيس كولنز و أشار الى البيانات الكثيرة التى تعارض التطور و تتنافى مع زعمه فى كتبه أن كل البيانات تدعم النظرية. ربما نحسن الظن و نقول أنه لا يقصد أن تجعل الدراونة يقرروا بالنيابة عنك بل يقصد أن تقرير الأمور يحتاج الى متخصصين و لكن احسان الظن يصبح صعبا جدا عندما ننظر الى كميات التضاربات و المشاكل فى أشجار الحياة التى لا يتم عرضها على الناس مقابل عرض جانب واحد فقط من البيانات منتقى بعناية يقول أن أشجار الحياة التطورية متسقة لكن العجيب أن قائل هذا الكلام فى المناقشات مع النقاد هو ذاته من يؤكد فى كتبه الموجهة للعامة ككتاب "انقاذ داروين" أن أدلة التطور ثابتة و واضحة ككروية الأرض
"Biologists today consider the common ancestry of all life a fact on par with the sphericity of the earth"
Karl Giberson "Saving Darwin: how to be a christian and believe in evolution" (2008): 53
Karl Giberson and Francis Collins "The language of Science and Faith" p.49
انقاذ داروين من ماذا بالضبط؟ هل يحتاج العلم المبرهن عليه بالأدلة الى انقاذ؟ أم أنها محاولة لانقاذ داروين من الأدلة التى يرفض جايبرسون و أمثاله عرضها على الناس و يكتفون بالأسلوب الكهنوتى المذكور فى الاقتباسات أعلاه "العلماء مؤمنون بالتطور" مع الأسف جايبرسون و شريكه فى كتابة الكتاب الثانى كولنز من المؤمنين و يمثلان نموذجا لكيف تمتطى خرافات المادية بعض المؤمنين لنشر نفسها بين العقلاء و صرفهم عن التفكير النقدى و استعراض الأدلة
يتم تصنيف الكائنات الحية وفقا لما يعرف بالتنادد homology و هو تشابه البنية بينهم حتى لو كانت هذه البنية تقوم بوظيفة مختلفة و قد اعتمد داروين على التنادد/التشابه كدليل انحدار الأنواع المختلفة من سلف مشترك. تخيل داروين شجرة الحياة بسلف مشترك فى الجذع تتفرع منه الكائنات الوسيطة حتى نصل الى الأنواع الحديثة على الأطراف بدون أى انقطاعات فهل يتوافق هذا الكلام مع سجل الحفريات؟
الاجابة و بمنتهى البساطة : لا. ان السمة الرئيسية للسجل الحفرى هى الظهور الانفجارى/المفاجئ للأنواع الجديدة و لعل أشهر هذه الانفجارات [وليس الوحيد] هو الانفجار الكمبرى حيث ظهرت الشعب الحيوانية phyla فى نفس الوقت تقريبا. كان داروين على علم بهذا سنة 1859 و أقر به فى كتابه ص 308 و لكنه كان يأمل أن تكتشف الحفريات الأعداد الضخمة المطلوبة من الحلقات الانتقالية و لكن بعد أكثر من 150 عاما من الحفر فى طبقات الأرض لا زال الدراونة لم يجدوا الحلقات الانتقالية الافتراضية. ان تصاميم و مخططات الأجساد المختلفة تتطلب ضخ قدر كبير من المعلومات الجديدة فى الحوض الجينى للكائنات الحية و المعلومات لا تنتجها العشوائية.
[يظن البعض أن الانفجار الكمبرى هو الشكل الانفجارى الوحيد فى السجل الأحفورى و هذا غير صحيح بل ان الظهور الانفجارى هو سمة السجل الأحفورى على طول الخط أيضا يظن البعض أن مجرد غياب الحلقات الوسيطة هو المشكلة بينما الحقيقة أن شجرة السجل الأحفورى هى عكس الشجرة التطورية بغض النظر عن الأشكال الانتقالية و للمزيد من التفاصيل حول الانفجارات الحفرية غير الكمبرية و تناقض الحفريات مع التطور يمكن مراجعة الفصل الخاص بالسجل الأحفورى و نفيه للداروينية من كتاب تصميم الحياة: t.ly/hg3z ]
التوازن المتقطع punctuated equilibrium
الى جانب مسألة الظهور الانفجارى فان السجل الحفرى حافل بالملاحظات المتناقضة مع التطور و من أهمها الثبات فمعظم الأنواع بعد أن تظهر فجأة تستمر بدون أن تتغير حتى تنقرض و هو الأمر الذى دفع التطوريين Stephen Jay Gould و Niles Eldredge الى وضع فرضية التوازن المتقطع لتبرير هذا التناقض بين الأحافير و التطور فافترض انعزال بعض الجماعات الصغيرة عن بقية النوع ثم تطفرها بسرعة لتتحول الى نوع جديد و بسبب صغر حجم الجماعة و سرعة التطور لا تترك الحلقات الانتقالية حفريات لنكتشفها. كلام لطيف جدا و لكنه فى الواقع عبارة عن طرح سيناريو غير مدعوم بأى أدلة فقط للتهرب من تعارض السجل الحفرى مع التطور. بل ان هذه الفرضية نفسها تتعارض مع اليات التطور التى تحتاج جماعات سكانية كبيرة و وقتا طويلا حتى تحدث الطفرات بأعداد ضخمة و حتى يكون هناك تنافس عالى بين الكائنات مما يقوى من دور الانتخاب الطبيعى
[و فى النهاية فان ستيفن جاى جولد واضع النظرية نفسه قد تراجع عنها و نقل هذا رتشارد دوكنزو سماه: استسلام جولد لداروين Gould’s Final capitulation to Darwin ]
الحفريات و تطور السلالات
لكن لنفترض جدلا أن لدينا حفريات كيف نثبت علاقة السلف بالأبناء؟ اذا كنا نرى بأعيننا اليوم تنوع سلالات الكائنات الحية المتعاصرة المنتمية لنفس النوع ألا يمكن أن تكون الحفريات هى لسلالات مختلفة من نفس النوع لكن عاشت فى أزمنة مختلفة؟ كيف يمكن علميا اثبات أن مجرد اختلاف عصر الحفرية معناه وجود علاقة سلف مع حفرية أخرى؟ برغم عدم قابلية هذه الفرضية للاثبات العلمى الا أننا نجد الأوراق العلمية التطورية مليئة بأشجار النسب بين الكائنات [يذكر لنا دكتور جيرى برجمان نموذج عملى لهذه المشكلة فى كتاب شهادة الأحافير Fossil Forensics : كمثال عملى لذلك فان الأسود و النمور لها هياكل متشابهة جدا فان وجدت فى الأحافير فقط كانت لتصنف على أنها نوع واحد و فى المقابل اذا وجدت الكلاب متباينة الأحجام كالشيواوا و الدانمركى و البلدج و الداشهند و الكولى فى الأحافير فقط فانها كانت ستصنف على أنها أنواع مختلفة برغم أن الحقيقة أنها نفس النوع
Mackenzie, Leslie, David K. Arwine, Edward J. Shewan and Michael J. McHugh. 2008. Biology: A Search for Order in Complexity. Arlington Hights, Il. Christian Liberty Press.
البطريق أيضا كان يمكن أن يكون حلقة انتقالية ممتازة لو لم يكن حيا لنعرف أن أجنحته هى فى الواقع زعانف عالية الكفاءة
من المشاكل الملفتة أيضا فى الكلادوجرام أو شجرة الحياة أن التفرع المفترض من السلف يكون مبنى على صفة مشتركة مع هذا السلف و لكن هناك صفات مختلفة تتشارك فيها كائنات مختلفة لذا فبمجرد تغيير الباحث للصفات التى بنى عليها التفرع تنتج لديه علاقات و تفرعات و أشجار مختلفة تماما
Wills, Matthew A. 2001. How good is the fossil record of arthropods? an assessment using the stratigraphic congruence of cladograms. Geological Journal 36: 206
Weygoldt, P. 1996. Evolutionary morphology of whip spiders: towards a phylogenetic system. Journal of zoological systematics and evolutionary research. 34:185-202
أى أن هناك عنصر غير موضوعى فى عملية التصنيف كما سبق التوضيح فى تفنيد أركان الداروينية t.ly/ZR4i ]
كيف تعمل أشجار التصنيف السلالى cladogram؟
"One cannot think of any observation which in case it would represent a true statement, would allow to conclude the falsity of a clade or a given cladogram through modus tollens"
L. Vogt "The unfalsifiabilitry of cladograms and its consequences" Cladistics 24(2008)
لا توجد أى صفة يمكن أن تجعلنا نعتبر رسما ما للعلاقات التطورية خاطئا
لارس فوجت - عالم نظم بيولوجية
التصنيف و التفسير:
بعيدا عن التصنيفات الشجرية الكبرى لشجرة الحياة ككل فكثيرا ما نرى تصنيفات شجرية "محلية" لمجموعات متقاربة و يتم عادة الاستدلال بمثل هذه الأشجار أو الكلادوجرام على انحدار هذه الأنواع من بعضها البعض و لكن دعنا نفهم أولا ما معنى أن يتم اعتبار كائن ما مرحلة سابقة على كائن اخر فى الأشجار التطورية اذ أن الترتيب بهذا الشكل عادة ما يعطى الانطباع بأن الكائن السابق هو "نسخة مخففة" watered down version أو نسخة أقل فى الصفات من الكائن التالى و بالتبعية فهو يكتسب صفات و يزداد تعقيدا و "يتطور" مع الوقت بغض النظر عن الالية و هذا الانطباع غير صحيح اذ أن الكائن السابق أحيانا تكون له صفات خاصة تميزه يتم تجاهلها تماما لمجرد أنها لا تربطه بالمجموعة تطوريا
“Cladistics, by contrast, gives no weight to unique characters”
Keynyn Brysse “From weird wonders to stem lineages: the second reclassification of the Burgess Shale Fauna” Studies in history and philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences 39, no. 3 (September 2008): 298 – 313
“The cladistic approach on the other hand, focuses on shared characters. Unique attributes, autopomorphies, are of no use in assessing relationship and are consequently accorded little significance”
Derek Briggs and Richard Fortey “The early radiation and relationships of the major arthropod groups” Science 246 (1989) : 241 – 243
باختصار شديد ما يفيد فى الترتيب التطورى للسلالات يتم أخذه و الباقى كأن لم يكن بناءا على ذلك يتم تقسيم الكائنات الى ما يسمى stem groups مجموعات جذع/ساق الشجرة و crown groups مجموعات تاج/قمة الشجرة بشكل يوحى بأن الساق بالضرورة يحتوى على صفات أقل كما و كيفا من التاج و هذا غير صحيح. لنأخذ مثالا عمليا على ذلك عند رسم شجرة التطور للمفصليات و تحديدا ثلاثيات الفصوص trilobite التى تعتبر من الكائنات المعقدة جدا التى تظهر فى الانفجار الكمبرى تم اعتبار مجموعة تسمى Radiodonta مجموعة سلف/جذع بسبب العثور على كائن Anomalocaris يمتلك عيون معقدة و لا يمتلك باقى الصفات
John R. Paterson et al., "Acute vision in the giant Cambrian predator Anomalocaris and the origin of compound eyes" Nature volume 480, pages237–240 (2011)
انظر الشكل رقم 3 فى الورقة
نفس هذا الكائن عند دراسته نجد أنه يمتلك صفاته الخاصة التى تميزه و لا يتم اعارتها انتباه لمجرد أنها غير مرتبطة تطوريا بالمجموعة التاج المطلوب تصنيفها/تفسيرها فى الشجرة مما يطعن فى امكان اعتبار التصنيف بهذا الشكل "تفسيرا" تطوريا
“Anomalocaris...is unlike any known arthropod, particularly in the nature of the jaw apparatus and the close-spaced, strongly overlapping lateral lobes”
Harry B. Whittington and Derek E.G. Briggs “The largest Cambrian animal, Anomalocaris, Burgess Shale, British Columbia” Philosophical Transactions of the Royal Society B 309 (1985): 569 – 609
لذلك فان شكل الشجرة اللطيف الذى تراه و تظن أنه كائنات تواصل اكتساب الصفات و التطور هو فى الواقع كائنات يتم تجميعها وفق الصفات المشتركة و تجاهل صفاتها الخاصة. بل و حتى فى الحالات التى تغيب فيها الكثير من الصفات من الأحفورة تعترف المراجعات العلمية أن سبب الغياب قد يكون التحلل و رداءة الحفظ لا عدم وجود هذه الصفات لذا فان مجموعات الساق كثيرا ما تصبح سلة لالقاء الكائنات رديئة الحفظ
A complete anatomy is never preserved and so it can be extremely difficult to know whether characters are absent because they have rotted away, or because they were not present to begin with.(72)...it will always remain difficult to differentiate stem members that lack crown group characters for taphonomic reasons from those that never had them. This problem is especially acute at deeper levels within a stem because, by definition, these organisms will share fewest characters with their respective crown groups and, at increasingly deeper levels in phylogenies, where homologies between component crown groups become increasingly obscure and less likely to be preserved (79,80)
Philip C. J. Donoghue and Mark A. Purnell "Distinguishing heat from light in debate over controversial fossils" Review article BioEssays 31:178–189
بحث اخر يوضح نفس المشكلة اذ يشرح كيف أن العظام المتصلة بالمنقار فى الطيور ان كانت متحركة تستخدم لتصنيف الفكيات الجديدة new jaws و غير المتحركة تستخدم لتصنيف الفكيات القديمة old jaws و التى تعتبر بقايا من الخطوط التطورية لأسلاف الفكيات الجديدة و يدعم هذا أن الديناصورات غير الطائرة التى يفترض كونها سلفا للطيور كانت من الفكيات القديمة ذات العظام المندمجة fused و لكن يتضح ان هناك طيورا قديمة جدا تمتلك الخاصية الأحدث
The bird - named Janavis finalidens - shares crucial features with its modern cousins...a bone called the pterygoid. “It is from a very interesting part of the skull, from the bony palate of the bird,” says Field. The bony palate has crucial features that researchers use to group birds, both living and extinct...the small, delicate pterygoid is usually missing from fossils
"67-million-year-old fossil upends bird evolutionary tree: Beak bone from ancient bird revises order of their modern ancestors." Nature 30 November 2022
لاحظ هنا كيف يقول أن عظمة مفصلية لعملية التصنيف و التى ينبنى عليها تصور الكائن عادة ما تكون غائبة فى الأحافير...يعيدنا هذا الى نفس ما قلناه ن تقسيم المجموعات الى ساق و تاج و أسلاف و أخلاف ينبنى على غياب صفات ربما تكون غائبة بسبب التحلل أصلا و ليس بسبب عدم وجودها فى الكائن ابتداءا...حسنا و ما التفسير؟ سهل جدا و بسيط: الصفة تطورت ثم فقدت و ان لم تعجبك الصيغة فيمكنك أن تقول أن الصفة القديمة تطورت تقاربيا أكثر من مرة بشكل مستقل و لا عزاء لاتساق الأشجار التطورية و الاستدلال بالصفات المشتركة على السلف المشترك
Its resemblance to the unfused pterygoids of modern chickens and ducks suggests that neognaths’ mobile beaks evolved first, and palaeognaths’ fused beaks arose later, says Field...the fused palate in modern palaeognaths could be a case of convergent evolution, says Field. Flightlessness evolved independently in emus and ostriches, which sit on different branches of the modern palaeognath tree, he says. It’s possible that those lineages also evolved fused palates independently.
المشكلة الثانية أنه حتى فى حالة هذه الصفات المشتركة فهى عادة غير موزعة عبر الشجرة بشكل "تطورى" يدعم فكرة الانحدار من سلف مشترك و اكتساب الصفات تدريجيا بل متناثرة بين الأنواع المختلفة و يقوم فريق البحث بتحديد الصفات التى يعتبرونها أهم و بناءا على ذلك يتم رسم الشجرة التطورية. و لنبنى على المثال السابق رأينا فى الشكل رقم 3 فى الورقة قبل السابقة تصنيف المفصليات Arthropods كمجموعة منحدرة من الشعاعيات Radiodonta – الان و لمجرد أن فريق اخر قرر أن الصفة الأهم من العين المركبة هى الأرجل المفصلية يتغير التصنيف (الذى يتم اعتباره شجرة تطور) فتصبح الشعاعيات مجموعة شقيقة للمفصليات و ليست سلفا و يصبح السلف هو فصيات الأقدام lobopods – انظر الشكل رقم 4 فى الورقة
Jianni Liu et al., "An armoured Cambrian lobopodian from China with arthropod-like appendages" Nature volume 470, pages526–530 (2011)
و طبعا تظهر نفس مشكلة الصفات الخاصة بالكائن و التى تميزه عن غيره و لا يتم اعتبارها لأنها ليست مهمة للتطور اذا تعترف نفس الورقة السابقة بأن الكائن له صفاته الاستثنائية و لكن نظرا لأنه امتلك صفة الأرجل المفصلية التى يعولون عليها فى تصنيف مفصليات الأرجل فربما اشتقت منه المفصليات لاحقا
Irrespective of its exact position, Diania, with its stout and spiny limbs attached to a slender, vermiform body, remains a highly unusual creature. It is hard to envisage it as the progenitor of any modern arthropod group, yet it may derive from a grade of lobopodian that acquired a key evolutionary innovation—and the name-giving character for Arthropoda—whereby sclerotized, jointed appendages began to fully develop.
ان المشكلة كما يقولون هى أن الصفات موزعة بين الكائنات بشكل mosaic يجعل كل كائن تركيبة خاصة من الصفات
However, we should caution that dinocaridids, Diania and other potential stem-arthropods typically express mosaics of arthropod-like characters, which makes resolving a single, simple tree of arthropod origins problematic.
و هو امر طبيعى ان توقفت عن الاصرار على حشر كل شئ فى قالب التطور الضيق فكل كائن توليفة خاصة من الصفات لأنها تلائم نمط حياته بدون أى داعى لفرضية السلف المشترك التى أربكت كل شئ و لعل من الأمثلة على ذلك أيضا معضلة توزيع صفات النمل بشكل غير تطورى بالمرة (حسب نمط حياة كل نوع) و التى ناقشناها فى هذا المقال فى المبحث المسمى: معضلة توزيع الصفات
لذلك يبدو واضحا أنه حتى مع محاولة التركيز على الصفات المشتركة فقط و تجاهل الصفات المميزة لكل كائن (و هذا خطأ فى حد ذاته) يظل بامكاننا انتاج عدة أشجار تطورية وفقا للصفة التى يظنها كل فريق أهم فمن ظن أن العيون المركبة أهم سينتج تسلسل و من ظن أن الأرجل المفصلية أهم سينتج تسلسل اخر و فى كلتا الحالتين يتم اللجوء للتطور المتقارب أو تطور الصفات ثم خسارتها ثم تطورها مجددا أو أى سيناريو اخر لتبرير وجود الصفات الأخرى التى لم يستخدمها فى تصنيفه فمن صنف وفق العيون المركبة سيضطر لافتراض نشأة الأرجل المفصلية أكثر من مرة و من صنف وفقا للأرجل المفصلية سيضطر الى افتراض نشأة العيون المركبة أكثر من مرة.
Key innovations include: 1 compound eyes (as figured for Odaraia), although this characterhas disappeared and ‘re-evolved’ a number of times with Arthropoda
David A. Legg et al., "Cambrian bivalved arthropod revealsorigin of arthrodization" Proceedings of the Royal Society B, Biological Sciences, 07 December 2012 Volume 279 Issue 1748
Legg et al.2 drew attention to the absence of sclerotized trunk limbs in dinocaridids (Anomalocaris, etc.), which remains for us one of the great puzzles of stem-group arthropod evolution. Put simply, dinocaridids have a more arthropod-like head region (cephalisation, eyes, sclerotized mouthparts), whereas Diania lacks such sophistication in the anterior body region, but has jointed trunk appendages. One solution to this puzzle would be to assume that dinocaridids also had trunk limbs, but lost them secondarily. In this scenario Diania could sit comfortably as sister group to a (Dinocaridida + Arthropoda) clade: all three sharing jointed appendages, the latter two sharing cephalisation, etc.
Jianni Liu et al., "Liu et al. reply" Nature volume 476, pageE1 (2011)
لذلك فان هذه الأشجار لا يمكن اعتبارها أبدا دليلا على التطور بل هى عملية تصنيف لا أكثر و هى فى الواقع تثبت أن الصفات موزعة عبر "شجرة الحياة" بشكل يتضارب مع التوقعات التطورية و فكرة السلف المشترك لأنه مهما كان يقينك من صحة الصفة التى اخترتها كأهم صفة للتصنيف ستظل حقيقة أن الصفات الأخرى نشأت بلا سلف مشترك و أن مجموعات الجذع لها صفات أخرى تميزها و قد علقت واحدة من أهم الخبراء فى تطور رباعيات الأقدام على هذا الأمر قائلة أنه بحسب الصفات المعتبرة و غير المعتبرة تتغير السيناريوهات التطورية
Depending on which characters were ordered versus unordered and which taxa were included or excluded, different results were obtained. In other words, the phylogeny of Devonian tetrapods was unstable
Clack, J. 2012. Gaining Ground: The Origin and Evolution of Tetrapods
نفس الأمر يتكرر فى واحدة من أشهر الأمثلة التطورية – الطيور اذ يجد التطوريون الصفات المطلوبة فى السلف موزعة بين نوعين من الديناصورات طيريات الورك Ornithischia لأسباب واضحة من الاسم و سحليات الورك Saurischia بسبب هيكل الحوض و طبعا نعلم جميعا أن كل هذه الديناصورات لها سماتها الخاصة فهى ليست مجرد نسخ مخففة من الطيور و يمكن مراجعة شهادة الأحافير: الطيران لرؤية تفاصيل أكثر عن تعدد السيناريوهات و الأشجار الممكنة و التى تتسق كلها مع البيانات. لذلك يعترف الكثير من خبراء التصنيف التطورى السلالى بأن طريقة انتقاء الصفات تؤدى الى ترك معلومات أخرى مهمة
“The complexity and number of potential phylogenetically informative characters often forced scientists to base their hypotheses on a few selected characters...leaving out much other important information and carrying the risk of producing biased results based on homoplasies”
Gregory D. Edgecombe et al., "Higher-level metazoan relationships: recent progress and remaining questions" Organisms Diversity & Evolution volume 11, pages151–172 (2011)
ان مشكلة التطوريون هنا عادة ما تكون كيف نحسم أى صفة هى القادمة من السلف المشترك و أيها هى التى نشأت بشكل مستقل لكن الحقيقة بالنظر الى أننا قادرون على رسم شجرة وفقا لأى سيناريو يفترض أى صفة فى السلف المشترك و الأخرى نشأت بشكل مستقل تصبح فرضية السلف المشترك قيمتها التفسيرية صفر لأن أى صفة يمكن أن تكون فى السلف المشترك و أى صفة يمكن أن تنشأ بشكل مستقل و بما أن أى صفة يمكن أن تكون نشأت بشكل مستقل اذن فكل صفة يمكن أن تكون نشأت بشكل مستقل لولا أنهم يحتاجون الى وضع سلف مشترك "قسرا" لأن نظرية التطور تملى عليهم ذلك. و من جديد بالعودة الى المثال السابق نقول ان كانت الأعين المركبة يمكن أن تنشأ بشكل مستقل أكثر من مرة و الأرجل المفصلية يمكن أن تنشأ بشكل مستقل أكثر من مرة فلماذا لا نفترض نشأة الصفتين بشكل مستقل فى المفصليات؟ السبب الوحيد هو أننا لو فعلنا ذلك سنلقى بنظرية التطور فى سلة القمامة لا أكثر و ليس أن البيانات أو عملية التصنيف تمنعه!!!
أيضا من المغالطات الشهيرة التى تقع فيها هذه الأشجار هى التعامل مع سلالات مختلفة و كأنها مراحل انتقالية نحو بعضها البعض فمثلا يتم اعتبار بعض أنواع الترايلوبايت مراحل انتقالية نحو بعضها البعض بينما كلها تنويعات من الترايلوبايت
“Early trilobites show all the features of the trilobite group as a whole, there do not seem to be any transitional or ancestral forms showing or combining the features of trilobites with other groups (e.g. early arthropods).”
P. Jell “Phylogeny of early cambrian trilobites” in Philip D. Lane et al “Trilobites and their relatives: Contributions from the third international conference” vol. 70 – London Palaeontological Association (2003): 45 -47
فهم يظهرون من البداية بصفاتهم المميزة كالعيون المركبة ثم يتم التنويع عليها لاحقا بل ان كثيرا من التنويعات نفسها تظهر بشكل انفجارى أيضا فى حادثة التشعب الأوردوفيشى العظيم (لاحظ أننا لا نقر التطوريين بأن مجرد وجود تنويعات على تصميم ما فانها قد حدثت لأسباب صدفوية مع انتخاب طبيعى و لكن الشاهد هو وجود أصل تلته تنوعات و بالتبعية هذه التنوعات ليست مراحل انتقالية بل تنوعات على تصميم أصلى سبقها - و من الطريف أيضا برغم الهوس التطورى بوضع سيناريوهات غير واقعية لامكان انتاج كل شئ بتراكمات تدريجية أنهم يقرون أن بعض العيون المركبة التى ظهرت لاحقا استعصت حتى على الخيال التطورى الواسع فى تصور امكان نشأتها تدريجيا )
They appeared close to the very beginning of the Cambrian Explosion of Life in the fossil record...and many groups of trilobites formed an important component of the Great Ordovician Diversification Event
The diversity of the morphology of trilobite eyes 'explodes' with the Ordovician....As such they are equipped from the very beginning of their appearance in the fossil record with elaborate compound eyes...We have seen that the general design of the holochroal eyes since the Ordovician, when the great ‘Biodiversification Event’ (GOBE) took place, changed from narrow, slit-formed eyes to compound eyes with wide visual fields and tunings to most different light situations. While all these adaptations can be achieved by modifications of the common principle of the holochroal eyes, the schizochroal eyes show up as not being apposition eyes. They seem to be a specialised, perhaps advanced sophisticated system, and are an object of current, continuing research.
Brigitte Schoenemann "An overview on trilobite eyes and their functioning" Arthropod Structure & Development Volume 61, March 2021, 101032
و نحن ان كنا نضرب المثال بالترايلوبايت هنا الا أننا ننبه أن هذه المشكلة شائعة فى كل التصانيف. مثال اخر على ذلك هو ما حدث عند مراجعة لتأريخ بعض الأحافير الخاصة بأسترالوبيثيكاس أفريكاناس الذى كان يفترض انحداره من أفارنسيس ليتضح أنه أقدم بكثير مما كان يظن و أنه معاصر لأفارنسيس ان لم يكن أقدم
Darryl E. Granger et al., "Cosmogenic nuclide dating of Australopithecus at Sterkfontein, South Africa" PNAS 119 (27)
"Fossils in the ‘Cradle of Humankind’ may be more than a million years older than previously thought" Purdue University (June 27, 2022)
حسنا و ما المشكلة اننا نحسن معرفتنا بكيفية حدوث التطور أليس كذلك؟ المشكلة أن "السهم" اللطيف الذى كان مرسوما من أفارنسيس الى أفريكانس و كان يتم اعتباره "دليل" قد تم محوه و بجرة قلم رسم سهم اخر فى مكان اخر على الشجرة و هذا هو ما نقوله مرارا و تكرارا. ان الأشجار التى يتصورها التطوريون و الأسهم التى يرسمونها ليست دليلا على شيئ و لا هى تثبت تحول نوع الى اخر بل هى مجرد تصورات ذهنية و نفس الصفات المورفولوجية التى تصلح كسلف يمكن أن تصلح كقريب معاصرلماذا؟ لأن هذه كلها قردة و التباينات بينها تباينات بين قردة و ليس بين كائنات تتطور من بعضها و حلقات انتقالية نحو بعضها و بالتالى فان الاستدلال بالأحافير و التصنيفات الشجرية محض توصيف للواقع و ليس دليلا فى حد ذاته. بجرة قلم سينتقل الأفريكانس من خلف لوسى الذى تطور منها الى الى فرع مجاور على الشجرة ليصبح أخ أو ابن عم تماما كما حدث مع اندوهيوس فى سلسلة الحيتان...أو ربما يتم ترجيح التأريخ القديم لأنه الأوقع بالنسبة للتطور كما حدث أيضا فى تطور الحيتان...و بعد كل هذا يصر التطورى على أن أشجاره دليل بل و تنبؤ و ليست مجرد تصنيف و توصيف!!!
الكارثة الأكبر أن تواريخ ظهور الكثير من الأنواع فى الأحافير كثيرا ما تتعارض مع أشجار الترتيب السلالى و هو ما يقول الى ممارسة تطورية عجيبة تعرف بالسلالات الشبحية
الأحفورة الشبح:
[اضافة هامة: من مقال دكتور بول نلسون - اشارة جمال الوزانى
من الممارسات العجيبة التى يمارسها الدراونة مفهموم السلالات الشبحية الذى استحدثه التطورى مارك نوريل. يشرح لنا نوريل فكرته قائلا: لنفترض أن هذا نموذج لجزءمن شجرة حياة مفترضة
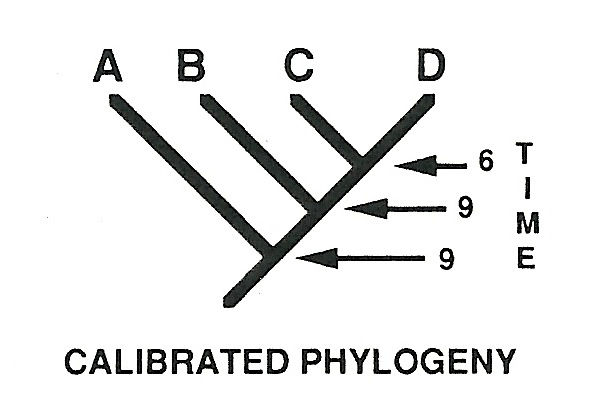
طبقا لهذا النموذج يجب أن تتفرع السلالات A وB أولا عند الوقت 9 (لنفترض مثلا منذ 9 مليون سنة) ثم تتفرع C وD بعدهما عند الزمن 6 ولكننا عندما نظرنا الى السجل الأحفورى وجدنا الاتى

كما نرى السجل الأحفورى يخالف توقعات شجرة الحياة فالسلالة A التى يفترض كونها بدائية قديمة تظهر حديثا جدا و السلالة D أيضا تظهر أحدث من توقعات شجرة الحياة. علميا معنى هذا أن شجرةا لحياة خطأ أما داروينيا فمعنى هذا أن السجل الأحفورى يحتوى على أشباح...نعم لا أمزح هكذا سماها التطوريون Ghost Lineage
Norell, Mark A. 1992. Taxic Origin and Temporal Diversity: The Effect of Phylogeny. In M.J. Novacek and Q.D. Wheeler, eds., Extinction and Phylogeny (New York: Columbia University Press), pp. 89-118.
يقول الدراونة المحترمون فى المصدر أعلاه طالما أن شجرة الحياة توقعت أن هذه السلالات يجب أن تكون موجودة فى حقب زمنية معينة فهى موجودة حتى لو لم نجدها فى السجل الأحفورى...هى ببساطة لم تحفظ ولكنها موجودة و كما يقول نوريل أسميها سلالات شبحية لأنها "خفية" على السجل الأحفورى و بالتالى يصبح شكله كما فى المصدر أعلاه هكذا

و باضافة مجموعة من الخطوط الحمراء تمثل السلالات الشبحية الخفية يتم حل المشكلة و التأكيد على أن السجل الأحفورى لا يتعارض مع الداروينية!!!
هل رأيتم؟ ان نظرية التطور تتحداكم لكى تثبتوا خطأها فقط أوجدوا أحفورة واحدة فى زمان غير ملائم ككائن يسبق سلفه. وجدتم؟ لا بأس السلالات الشبحية موجودة و سنفترض أنه كان موجودا لكننا لم نعثر عليه و لكننا لازلنا نظرية علمية قابلة للاختبار علميا...فقط عليكم أن تجدوا كائنا يسبق سلفه
هكذا اذن-شجرة الحياة التى ترسمها الداروينية صحيحة و على السجل الأحفورى أن "يحترم نفسه" و يوفق أوضاعه وفق هذه الشجرة. ان أى نظرية علمية تقدم تنبؤات عن الواقع و يتضح عدم صحتها تتلقى ضربة قوية أما الداروينية فهى النظرية الوحيدة التى يتم "تغيير الواقع و الأدلة" لتتماشى مع صحتها!!!]
مثلا بالعودة الى مثال المفصليات و الترايلوبايت السابق سنرى شيئا لطيفا جدا...شجرة التطور السلالى للمفصليات المرتبة بشكل أنيق تبدو تماما كما يقول التطور...و لكن ليس تماما كما تقول الأحافير اذ لا تظهر فيها المجموعات أصلا بالترتيب الذى تظهر به فى الأحافير بل بالترتيب الذى يحتاجه التطور فبعض الكائنات التى تمتلك صفة من صفات المفصليات دون الباقى لذا يفترض أنها سلف للمفصليات تظهر فى الأحافير بعد ظهور المفصليات.

Gregory D.Edgecombe "Arthropod phylogeny: An overview from the perspectives of morphology, molecular data and the fossil record" Arthropod Structure & Development Volume 39, Issues 2–3, March–May 2010, Pages 74-87
الخطوط السميكة هى ما يقوله السجل الأحفورى أما الرفيعة فهى سلالات شبحية – مجرد افتراض من التطوريين أن هذه الكائنات لابد أنها كانت موجودة و لم نكتشفها لأن التطور يتطلب أن تكون موجودة! بالنظر الى هذه الخطوط يظهر لنا أن الواقع يقول أن مجموعات الجذع تظهر مع أو بعد مجموعات التاج عكس ما يقوله التطور تماما بل انه من المثير للسخرية أن بعض أكثر الأسلاف المفترضة بدائية هى من اخر الكائنات ظهورا
و فى بحث اخر نجد نفس المشكلة يتم تصنيف كائن يظهر فى المرحلة الخامسة من الانفجار الكمبرى كسلف و مجموعة جذع للمفصليات بينما مجموعة التاج تظهر قبله فى المرحلة الثالثة كما فى الشكل أعلاه و الحل سلالة شبحية طبعا
Herein, we describe a new bivalved arthropod from the Tulip Beds of the Burgess Shale Formation (Cambrian, stage 5).
David A. Legg et al., "Cambrian bivalved arthropod revealsorigin of arthrodization" Proceedings of the Royal Society B, Biological Sciences, 07 December 2012 Volume 279 Issue 1748. See Figure 3, location of Nereocaris
و طبعا كالعادة عندما نجد أنه حتى بعد السلالة الشبحية لازالت لدينا مشكلة فى ترتيب ظهور الصفات من السهل أن نضيف فرضيات جديدة مثل الصفات تظهر و تختفى و تظهر مرة أخرى
Key innovations include: 1 compound eyes (as figured for Odaraia), although this character has disappeared and ‘re-evolved’ a number of times with Arthropoda

Philip C. J. Donoghue and Mark A. Purnell "Distinguishing heat from light in debate over controversial fossils" Review article BioEssays 31:178–189
ثنائية التناظر جاءت ثنائية التناظر ذهبت...الخياشيم جاءت الخياشيم ذهبت...ان أقوى أدلة التطور كما يقول رتشارد دوكنز هى الترتيب الشجرى التراكمى المثالى للصفات...واضح!!! الكارثة أن جمل مثل "الحصول على المخطط الجسدى الفلانى أو فقده”-حتى اذا سلمنا جدلا بحدوثها بالترتيب المذكور-تتطلب اعادة تنظيم جينية كبرى و اعادة ترتيب للأنسجة و الأعضاء و الاليات النمائية و لا يمكن افتراض حدوثها بهذه البساطة لمجرد أنها مرصودة فى كائن ما. و فى النهاية تجد هذه الأبحاث تصل و بمنتهى الجزم الى استنتاجات مفادها أن لدينا نماذج واضحة على التدرج و التطور...يذكرنا هذا بكلام ستيفن جاى جولد عالم الأحافير التطورى الشهير عندما قال أن أشجار التطور التى تملأ الكتب تحتوى على بيانات عند الأطراف و نقاط التفرع و لكنها لا تحتوى على بيانات كيف تنشأ صفة انتقاليا بل ان بعض الحالات لا يمكن أصلا تصور ذهنيا كيف تنشأ مثل هذه الصفة تدريجيا و حالات أخرى تم وضع التصور على هيئة قصة غير مثبتة
“The evolutionary trees that adorn our textbooks have data only at the tips and nodes of their branches; the rest is inference, however reasonable, not the evidence of fossils...In any local area, a species does not arise gradually by the steady transformation of its ancestors; it appears all at once and ‘fully formed.’”
Gould, Stephen Jay. “Evolution’s erratic pace.” Natural History 86 (May, 1977): 12 - 16
“The absence of fossil evidence for intermediary stages between major transitions in organic design, indeed our inability, even in our imagination, to construct functional intermediates in many cases, has been a persistent and nagging problem for gradualistic accounts of evolution.”
Gould, Stephen Jay. “Is a new and general theory of evolution emerging?” Paleobiology 6(1): 119-130 (1980)
Two other classes of information were explained away or simply ignored: 1) morphological gaps in stratigraphic sequences...2) evolutionary stasis,...the gradualistic idols that were established had feet of clay and rarely survived an intensive restudy...The collapse of classic after classic should have brought these gradualistic biases into question. The alienation of practical stratigraphy from an evolutionary science that required gradualism should have suggested trouble
Gould, Stephen Jay and Niles Eldredge. “Punctuated Equilibria: The Tempo and Mode of Evolution Reconsidered.” Paleobiology 3 (2) (Spring, 1977): 115-151.
"the majority of well documented examples...had never been adequately explained in the terms demanded by Darwinian convention-that is, as adaptive improvements of constituent organisms in anagenetic sequences. Most so-called explanations amounted to little more than what Lewontin and I, following Kipling, would later call “just-so stories,” or plausible claims without tested evidence, whereas other prominent trends couldn’t even generate a plausible story in adaptationist terms at all."
Gould, Stephen Jay. The Structure of Evolutionary Theory. Cambridge, MA: Belknapp Press of Harvard University Press, 2002. PP. 38-39
ان هذه النماذج لم تصبح واضحة و متوافقة مع التطور الا بعد اضافة كل هذه الافتراضات كالسلالات الشبحية و تطور الصفات أكثر من مرة و تجاهل الصفات المميزة للأنواع و التركيز على القواسم المشتركة فقط

أشجار التطور المفترضة للثدييات و الطيور: الخطوط السوداء السميكة فى النصف الأعلى فقط هى ما يمكن اعتباره حقيقة و الباقى افتراضات من كيس نظرية التطور أى أن النصف الأسفل من الشجرة بالكامل وهم غير موجود فى الحقيقة. لماذا يفترضون هذا؟ لأن النظرية تحتاج الى وقت حتى تتشعب فيه هذه الكائنات من سلفها المفترض بالتدريج خاصة و أنهم يعترفون أن الفروق الجينية بينها كبيرة surprisingly extensive و الكائنات تظهر فجأة متمايزة و متباينة عن بعضها بدون تشعبات من سلف مشترك تسبقها اذن فبدلا من افتراض أن النظرية خطأ يتم افتراض أن البيانات هى الخطأ لأنها لا تلائم النظرية و يتم وضع خيالات غير موجودة فى الواقع لسلسلة التطور و التشعب. (ملحوظة: المزاعم بشأن ضعف السجل الأحفورى أو التطور السريع المسمى التوازن المتقطع تم الرد عليها سابقا عند الحديث عن الأحافير و توضيح أنها فرضيات من كيس النظرية لتبرير تعارض البيانات معها)
Cladistic and biogeographical data indicate the Early Tertiary ‘explosion’ of mammalian and avian diversity was preceded by a period of unobserved diversification, which molecular data suggests was surprisingly extensive. Thick solid lines: known range. Shaded lines: ghost range implied cladistically.
Alan Cooper and Richard Fortey "Evolutionary explosions and the phylogenetic fuse" Trends in Ecology and Evolution REVIEW ARTICLE VOLUME 13, ISSUE 4, P151-156
و من الأمثلة الشهيرة أيضا على ذلك أنه فى العصر الديفونى تظهر أولى اثار أقدام رباعيات الأقدام البرية قبل ظهور أسلافها المفترضين من الأسماك فى الأحافير و هو ما يتعارض مع خط التطور الذى يشيعه التطوريون
Grzegorz Niedźwiedzki et al., "Tetrapod trackways from the early Middle Devonian period of Poland" Nature volume 463, pages43–48 (2010)
"The temporal mismatch implies the existence of long 'ghost ranges' (dashed red lines) among Devonian tetrapodomorphs."
Philippe Janvier & Gaël Clément "Muddy tetrapod origins" Nature volume 463, pages40–41 (2010)
لاحظ الشكل الموجود بالورقة الأخيرة Figure 1 لتدرك كيف يتعامل التطوريون مع البيانات...بمنتهى البساطة افترضوا وجود ghost range سلالة شبحية مشار اليها بالخط الأحمر المنقط لابد أنها كانت موجودة و لكن لم يعثر عليه. أيضا نقاط التفرع التى ينحدر منها نوعان او أكثر و لا يكون محددا لها اسم عادة ما تكون "كائن افتراضى" يفترضون وجوده لأن التطور يحتاج اليه
البيولوجيا الجزيئية:
“عندما نقارن التسلسلات الجينية بين المخلوقات المختلفة سنجد نفس الترتيب الشجرى التى نحصل عليها عند مقارنة التشابهات التشريحية"
رتشارد دوكنز "أعظم استعباط على وجه الأرض: أدلة التطور" ص 315
“ان الجينات تؤكد نفس العلاقات التطورية التى تنتج من التحليل التشريحى"
جيرى كوين "لماذا التطور حقيقة" 10
“ليس هناك أمثلة على خرق هذه المشاهدات [توافق الشجرة التشريحية مع الجزيئية]”
بيتر أتكنز "اصبع جاليليو: الأفكار العشرة العظيمة فى العلم" ص 16
“[ان البيانات التشريحية و الجزيئية] تتقارب نحو اجابة واحدة بأوكد ما يكون"
دونالد بروثيرو "التطور" ص 140
لحظة واحدة أيها السادة!!! من المفترض وفقا لأسس علم الأحياء أن الحمض النووى يحمل شفرة بناء الكائن الحى و بالتبعية فالكائنات ذات السمات التشريحية المتشابهة يجب أن تحمل شفرة متشابهة و كلما ابتعدت هذه الكائنات عن بعضها تشريحيا فيجب أن تتباعد شفرات بناءها فأين هو الدليل الاضافى الذى تم اكتشافه بالضبط حتى ندعى وجود دليل تشريحى و اخر جينى و هما يتلاقيان عند نفس الاجابة؟ أم أنها مجرد محاولة للايحاء بكثرة الأدلة؟ يشبه الأمر أن يقول لك أحدهم أنه عندما يقارن بين خصائص أنظمة تشغيل حاسب الى مختلفة فانه يجد بينها درجات من التشابهات و الفروق ثم-ويا للعجب-عندما يقارن بين كود/شفرة برامج التشغيل فانه يجدها تعكس هذه التشابهات و الفروق و هذه مسألة مستقلة تماما ليصبح عمل نظام التشغيل دليل و برمجة نظام التشغيل التى تقود الى هذا العمل دليل مستقل!!! هل تمزح؟!!! هل من المفترض أن تنظر الى برمجة نظام التشغيل فى جهاز يعمل بنظام ويندوز 11 لتجد الكود-و يا للمفاجأة-هو كود وينوز 7 مثلا أو لتجده أقرب الى كود ويندوز 2000 منه الى كود ويندوز 10 حتى تتظاهر بأننا من المفترض أن نعتبر هذا دليلا مستقلا فى حد ذاته؟ أم أنها مجرد محاولة رخيصة للايحاء بكثرة الأدلة بالخدع الكلامية التى يبرع فيها مروجو التطور؟ كأنك من المفترض أن تنظر الى مبنيين متشابهين بنسبة كبيرة ثم تنظر الى تصميمهما على الورق فتندهش لأن التصميمين متشابهين أيضا...ان هذه الأساليب الكلامية الخداعية التى تحاول خلق جو درامى يوحى للمستمع من غير المتخصصين بأن الأدلة كثيرة هى عين ما علق عليه المحامى الشهير فيليب جونسون كاتب كتاب "داروين فى المحكمة" Darwin on Trial قائلا أنها ما يستخدمه بعض المحامين من أساليب خطابية للتعمية على ضعف أدلة موقفهم [لا أعرف لماذا يذكرنى هذا "الدليل" بجملة فى مسرحية شهيرة تسمى "شاهد مشفش حاجة" عندما قال أحدهم: “انها راقصة...و الى جانب ذلك لدى اضافة مهمة...انها ترقص!!! فعلا انها اضافة مهمة و غير متوقعة بالمرة و مفاجأة و دليل اضافى على أنها راقصة!!! انه “دليل مدلش على الحاجة”]
و لكن هل تعرف ما هو أطرف شئ فى هذه المسألة؟ أن الكتاب السابقين و أمثالهم ليسوا مجموعة من السذج كبطل المسرحية المذكورة و لكنهم يكذبون...و بمنتهى الصفاقة. يقال ان أفضل طريقة للكذب هى أن تقول كذبة كبيرة جدا و تقولها بثقة شديدة و تصر عليها و عندها لن يتخيل المتلقى أبدا أن لديك الجرأة على كذبة بهذا الحجم بهذه الثقة و هذا الاصرار فسيفترض أنك صادق. و سنستعرض أدلة ذلك. مع التقدم العلمى و ظهور البيولوجيا الجزيئية التى تدرس تركيب الحمض النووى DNA [قاعدة البيانات المشفرة التى تخزن تصميم الكائن الحى] و البروتينات بدأ التطوريون محاولات استخدامها لبناء شجرة الحياة. [فمثلا لو وجدنا جينا فى نوع من الكائنات و نفس الجين فى نوع اخر فهذا نظريا دليل سلف مشترك بناءا على هذه الجينات المتماثلة Orthologous Genes فماذا كانت النتيجة؟ تم تدمير فرضية وجود شجرة تتشعب فيها الحياة تماما:
the history of life cannot properly be represented as a tree
W. Ford Doolittle, “Phylogenetic Classification and the Universal Tree,” Science, Vol. 284:2124-2128
Phylogenetic incongruities [conflicts] can be seen everywhere in the universal tree, from its root to the major branchings within and among the various taxa to the makeup of the primary groupings themselves
Carl Woese “The Universal Ancestor,” Proceedings of the National Academy of Sciences USA, Vol. 95:6854-9859 (June, 1998)
Wolf, Y. I., Rogozin, I. B., Grishin, N. V.,& Koonin, E. V. (2002). Genome trees and the tree of life. TRENDS in Genetics, 18(9), 472 – 479
Bapteste, E., Susko, E., Leigh, J., MacLeod, D., Charlebois, R. L.,& Doolittle, W. F. (2005). Do orthologous gene phylogenies really support tree - thinking?. BMC Evolutionary Biology, 5(1), 33.
Koonin, E. V., & Wolf, Y. I. (2010). The common ancestry of life. Biol Direct, 5(1), 64 – 64.
Leigh et al., 2011, Evaluating Phylogenetic Congruence in the Post-Genomic Era, Genome Biology and Evolution, Vol. 3, pp 571-587
فلو رسمنا علاقة بين الكائنات الحيّة بواسطة الجينات؛ فلن نجد أيّ شجرة، ولن نجد أى فروعً تتفرّع بعضها من بعض، بل سنجد البينانت تتضارب وفق الخصائص التى تم اختيارها للمقارنة وهذا يعدّ نقضًا لفرضيّة التّطوّر وتنبؤاتها
أحمد ابراهيم – اختراق عقل]
فى 2005 قام علماء البيولوجيا التطورية بمحاولة بناء شجرة تطورية بتحليل جينات مختلفة للكائنات الحية فجاءت النتائج متناقضة مع بعضها حسب الجينات المستخدمة
Antonis Rokas, Dirk Kruger, and Sean B. Carroll, "Animal evolution and the molecular signature of radiations compressed in time,” Science 310 (2005): 1933—1938. doi:10.1126/ science.1116759. PMID:16373569.ثم فى 2008 تمت محاولة أخرى لبناء شجرة تطورية جينية لبعض المجموعات الحيوانية
Casey W. Dunn, Andreas Hejnol, David Q. Mams, Kevin Pang, William E. Browne, Stephen A. Smith, Elaine Seaver, Greg W. Rouse, Matthias Obst, Gregory D. Edgecombe, Martin V. Sorensen, Steven H. D. Haddock, Andreas Schmidt-Rhaesa, Akiko Okusu, Reinhardt Mobjerg Kristensen, Ward C. Wheeler, Mark Q. Martindale, and Gonzalo Giribet, "Broad phylogenomic sampling improves resolution of the animal tree of life," Nature 452 (2008): 745-749. doi:10.1038/nature06614. PMID:18322464. فتعارضت مع شجرة أخرى تم بناءها فى عام تالى باستخدام جينات أخرى
Herve Philippe, Romain Derelle, Philippe Lopez, Kerstin Pick, Carole Borchiellini, Nicole Boury-Esnault, Jean Vacelet, Emmanuelle Renard, Evelyn Houliston, Eric Queinnec, Corinne Da Silva, Patrick Wincker, Herve Le Guyader, Sally Leys, Daniel J. Jackson, Fabian Schreiber, Dirk Erpenbeck, Burkhard Morgenstern, Gert Worheide, and Michael Manuel, "Phylogenomics revives traditional views on deep animal relationships,” Current Biology 19 (2009): 706—712. doi:10.1016/j.cub.2009.02.052. PMID:19345102.[الان أرجو منك أن تدقق فى العناوين علوم الجينات وجدت تشابهات بين الكائنات – علوم الوراثة تدعم التطور ما لا يقوله لك الدراونة المحترمون و لن تعرفه بمجرد قراءة العناوين بل بمقارنة الأبحاث ببعضها أن مستويات التشابه بين الكائنات (وبالتبعية علاقة السلف المشترك المفترضة) تختلف باختلاف البروتينات أو الجينات المستخدمة فى الدراسة و تختلف عن الشجرة المبنية على أساس التشابه الشكلى. ان هذه النتائج تتعارض بصورة كلية مع فرضية السلف المشترك و تدعم فرضية "المصدر المشترك" أو "التصميم المشترك" و الذى يتم فيه اعادة استخدام المكونات المناسبة فى المكان المناسب بغض النظر عن تباعد أو تقارب الكائنات]
[لقد قارن أستاذ علم الجينات بجامعة هارفارد مئات الجينات المشتركة بين العديد من الكائنات من بشر و ضفادع و قنافذ و ذباب فاكهة و ديدان و غيرها و كانت النتيجة أن الجينات تقدم قصصا تطورية مختلفة
"The problem was that different genes told contradictory evolutionary stories...it's not a tree anymore"
"the holy grail was to build a tree of life...today the project lies in tatters torm to pieces by an onslaught of negative evidence...[many] biologists now argue that the tree concept is obsolete and needs to be discarded"
الجينات المختلفة تعطى سيناريوهات تطورية مختلفة...انها ليست شجرة...الأدلة السلبية كثيرة جدا...ان مفهوم الشجرة لم يعد صالحا و يجب تركه
Graham Lawton, "Why Darwin was wrong about the tree of life, " New Scientist (January 21, 2009).
سامى عامرى - براهين وجود الله]
و هى نفس النتيجة التى توصلت اليها محاولات رسم شجرة تطور جينية للطيور اذ كلما ركزوا على جزء من الجينوم أعطاهم نتائج غير الجزء الاخر و النتائج لا تتفق مع الشجرة الأصلية المفترضة
While preparing their paper, Gilbert and his team had fiddled with their data set to understand the differences between gene trees. When they told their tree-building software to focus only on regions of the genome that Prum’s team used, it produced a tree that looked like Prum’s. When they shifted focus to other regions, a very different tree emerged. When they divided their bird genomes into thousands of different parts and ran each through their software, they got thousands of different trees, and not one completely matched the “species tree” they had constructed from large portions of genomes.
"The Bizarre Bird That’s Breaking the Tree of Life" The New Yorker July 15, 2022
All estimates of gene trees differ from our candidate species trees. No single intron, exon, or UCE locus from our TENT data set had an estimated topology identical to the ExaML TENT or MP-EST* TENT.
ERICH D. JARVIS et al., "Whole-genome analyses resolve early branches in the tree of life of modern birds" SCIENCE 12 Dec 2014 Vol 346, Issue 6215 pp. 1320-1331
و طبعا جميع أنواع الترقيعات التى يمكن تخيلها تقال من نقص البيانات الى تزاوج بين الأنواع المختلفة الى التطور السريع لكن السلف المشترك الذى تعارضه كل البيانات يجب الحفاظ عليه...الدراسات الجزيئية تتضارب مع المورفولوجية و مع بعضها البعض...الأصابع شبه الزوجية التى كنا نرسمه كسلف للأصابع الزوجية اتضح أنها تنشأ بشكل مستقل...الصفات تتطور و تفقد تتطور و تفقد...المجد للمرونة التطورية و لا تنسى أن لدينا أشجار انسيابية تدرجية رائعة
recent molecular studies have yielded topologies that conflict with those based on morphology and with one another...the morphology-based tree conflicts with recent molecular results.
independent evolution of semizygodactyly in Coliiformes, Leptosomiformes, and Strigiformes, none of which is closely related to extant clades exhibiting full zygodactyly... paleontological data instead support a scenario in which semizygodactyly arises independently...the fully zygodactyl Psittaciformes and Piciformes appear to share no close phylogenetic relationships with any of the three semizygodactyl clades.
Interestingly, a complete loss of zygodactyly is supported in our results; crown Passeriformes are strongly supported as the sister taxon to the extinct, fully zygodactyl clade Zygodactylidae, in agreement with recent work (42), and our character state reconstructions are consistent with the previously hypothesized scenario in which songbirds and parrots share a fully zygodactyl ancestor, with subsequent reversion to an anisodactyl condition in the lineage leading to modern songbirds. The numerous independent shifts in foot morphology inferred here reinforce our appreciation of the plasticity of digital conformation, which has been linked to embryonic development and muscle activity
Daniel T. Ksepka et al., "Early Paleocene landbird supports rapid phylogenetic and morphological diversification of crown birds after the K–Pg mass extinction" PNAS Vol. 114 | No. 30 July 10, 2017 114 (30) 8047-8052
اذا صنعت لك البيانات شجرة فان هذا دليل على التطور و اذا لم تصنع فهو أيضا دليل على التطور و سيتم حشر البيانات فى شجرة قسرا...لابد انها تطورت ثم فقدت أو تطورت أكثر من مرة...أى شيء الهم السلف المشترك حقيقة علمية
Sponges (Porifera) and placozoans lack neurons, so if comb jellies are a sister taxon to all other metazoans, then either those two taxa have lost neurons during evolution or neurons (and nerve nets) evolved twice independently (see Figure 3)... In contrast, comparative morphology and recent advances in solving animal relationships with molecular tools suggest that internalizations from a basiepidermal to a subepidermal condensation happened multiple times independently, for example inside the ribbon worms (Nemertea) and segmented worms (Annelida).
Andreas Hejnol and Fabian Rentzsch "Neural nets" Current Biology VOLUME 25, ISSUE 18, PR782-R786, SEPTEMBER 21, 2015
بل ان نفس الدراسة أحيانا قد تأتى بنتائج مختلفة حسب المقارنة المستخدمة. انظر مثلا شكل واحد و اثنان فى هذه الورقة لترى علاقات تطورية مختلفة بين نفس المجموعتان Rheas/Tinamous
Harshman J et al. 2008. Phylogenomic evidence for multiple losses of flight in ratite birds. PNAS 105(36), 13462–13467.
Smith JV, Brau EL, Kimball RT 2013. Ratite Nonmonophyly: Independent Evidence from 40 Novel Loci. Systematic Biology 62(1), 35–49.
و عندما أراد شون كارول فى كتابه الاستدلال على السلف المشترك بوحدة جين Pax-6 الذى يعتبر مفتاح تشغيل لعملية نمو العين اعترف ضمنيا بأن هذا يتعارض مع شجرة تطور العيون التى كانت مرسومة على أساس تشريحى
The great evolutionary biologist Ernst Mayr and his colleague L. V. Salvini-Plawen suggested, on the basis of the cellular anatomy of animal eyes, that eyes had been invented independently some forty to sixty-five times...Since Pax-6 is involved in eye development in such a wide range of animals, it is very unlikely that each happened upon the use of Pax-6 by accident. The widespread role of Pax-6 in eye development must be due to historical reasons. That is, a common ancestor of these animals used Pax-6 in the development of some, perhaps very primitive eye.
Sean B. Carroll, The Making of the Fittest: DNA and the Ultimate Forensic Record of Evolution p.194-196
تطور النباتات: جينات مختلفة تساوى أشجار مختلفة لنصل الى صورة لا علاقة لها بالمزاعم العريضة للنظرية

Andre S. Chanderbali et al., "Buxus and Tetracentron genomes help resolve eudicot genome history" Nature Communications volume 13, Article number: 643 (2022)
مثال اخر فى 2013 تمت مقارنة 10 جينات فى نحو 500 نوع من الأسماك لدراسة شجرة تطورها و كالعادة جاءت النتائج مربكة للأشجار المورفولوجية
Near, T and 10 other authrors. 2013. Phylogeny and tempo of diversification in the superradiation of spinyrayed fishes. Proceedings of the National Academy of Science. 110(31): 12738 - 12743
و نفس الأمر فى تصنيف الرخويات اذ كان الظن أن الاسفجيات مثلا أكثر بدائية و تفرعت مبكرا ثم تلاها ما هو أعقد بناءا على الصفات المورفولوجية ثم جاءت دراسات جينية بعكس النتيجة لترجح أنها فى الواقع كائن متدهور من سلف أكثر تعقيدا
"The Closest Living Relative of the First Animal Has Finally Been Found" Scientific American (May 17, 2023)
و أمثلة أخرى على تضارب الاستنتاجات المورفولوجية مع الجينية و الجزيئية
Evolutionary trees constructed by studying biological molecules often don’t resemble those drawn up from morphology
Trisha Gura, “Bones, Molecules or Both?,” Nature, Vol. 406:230-233 (July 20, 2000)
that molecular evidence typically squares with morphological patterns is a view held by many biologists, but interestingly, by relatively few systematists. Most of the latter know that the two lines of evidence may often be incongruent.
Masami Hasegawa, Jun Adachi, Michel C. Milinkovitch, “Novel Phylogeny of Whales Supported by Total Molecular Evidence,” Journal of Molecular Evolution, Vol. 44, pgs. S117-S120 (Supplement 1, 1997)
“the wealth of competing morphological, as well as molecular proposals [of] the prevailing phylogenies of the mammalian orders would reduce [the mammalian tree] to an unresolved bush, the only consistent clade probably being the grouping of elephants and sea cows
W. W. De Jong, “Molecules remodel the mammalian tree,” Trends in Ecology and Evolution, Vol 13(7), pgs. 270-274 (July 7, 1998)
Nardi, Francesco; Giacomo Spinsanti, Jeffery L. Boore, Antonio Carapelli, Romano Dallai, and Francesco Fratti. 2003. "Hexapod Origins: Monophyletic or Paraphyletic?" Science, 299: 1887-1889
Maley and Marshall "The coming age of molecular systematics" p.505
Rokas et al., "Conflicting phylogenetic signals at the base of the metazoan tree"
Halanych "The new view of abimal phylogeny"
Ronald Jenner "Higher level Crustacean phylogeny" p.143 - 151
Holton and Pisani "Deep Genomic-Scale analyses of the metazoa reject Coelomata"
Elie Dolgin, “Rewriting Evolution,” Nature, 486: 460-462 (June 28, 2012)
The researchers compared the genomes of two kinds of marine worms with simple morphology – Xenoturbellida and Acoelomorpha – with those of other animals. They demonstrated that their previous position at the base of the bilateral symmetry animal group – that includes insects, mollusks and vertebrates – was inaccurate. "Instead, we determined that Xenoturbellida and Acoelomorpha are closely related to the complex deuterostomes, which is a major lineage containing sea urchins, humans and sharks,"
"New evolutionary research disproves living missing link theories" EurekAlert 10-FEB-2011
"For over a hundred years, we've been classifying organisms according to how they look and are put together anatomically, but molecular data often tells us a rather different story."
"Study suggests that most of our evolutionary trees could be wrong" Phys.org (June 2022)
حتى المقارنات بين الأصابع الثلاث للطيور و الديناصورات الطائرة التى يفترض انحدارها منها فى واحدة من أشهر القصص التطورية أدت الى نتائج متضاربة بين الشكل و الجينات و الاليات النمائية
Xing Xu and Susan Mackem "Tracing the evolution of avian wing digits" Current Biology 2013 Jun 17;23(12):R538-44
كما تضاربت أيضا فرضيات أصول أجنحة الحشرات عند مقارنة السمات الشكلية المورفولوجية مع الاليات النمائية الجينية
David E. Alexander "On the Wing: Insects, Pterosaurs, Birds, Bats and the Evolution of Animal Flight" (2015)
بل حتى فى نطاق السمات التشريحية ذاتها و التى تعتبر أساس بناء الشجرة التطورية المعيارية تعترف المراجعات بأن العلاقات التطورية يمكن أن تتضارب وفق السمات التشريحية المستخدمة مثل تناظر الجسد symmetry و التجويف الجسدى coelom و أقسام الجسم segmentation و غيرها لتنتج أشجار متضاربة معا
Willmer and Holland “Modern approaches to metazoan rlationships” p. 690 – 691
Jenner “Evolution of animal body plans” p.209
3 أشجار متضاربة و مل واحدة لها من البيانات ما يدعمها بقوة
patterns of extreme inconsistencies at the base of the metazoan tree of life...Different gene matrices tell different stories...major conflicts between the previously published metazoan phylogenies...To summarize, this study generated three incongruent, yet strongly supported tree topologies
Nosenko et al., “Deep metazoan phylogeny: When different genes tell different stories,” Molecular Phylogenetics and Evolution, 67: 223-233 (2013).
و تشير الكثير من الأبحاث الى نفس النتائج كما يقر عدد متزايد من أقطاب البيولوجيا التطورية مثل يوجين كونين و اليزابث بنسى بأن جينومات الكائنات الحية لا يمكن تصنيفها كشجرة...طبعا يتم تقديم تفسيرات من طراز تبادل الكائنات للجينات عن طريق تهجين الأنواع المختلفة أو الانتقال الأفقى للجينات عن طريق البكتيريا و بغض النظر عن لامعقولية بعض التفاسير الا أن النتيجة النهائية هى أن صور الأشجار اللطيفة التى يتم القاءها يمينا و يسارا عند الاستدلال على التطور لم يعد العلم يدعمها (هذا اذا فرضنا أصلا أن مجرد ترتيب الكائنات على شكل شجرة يكفى لاثبات تحول أحدها الى الاخر)
In fact, conflicting phylogenetic signal between genes is commonplace.
Dominik Schrempf and Gergely Szöllősi (2020). The Sources of Phylogenetic Conflicts. In Scornavacca, C., Delsuc, F., and Galtier, N., editors, Phylogenetics in the Genomic Era, chapter No. 3.1, pp. 3.1:103.1:23 (2021)
An immediate challenge is to address the pervasive phylogenetic conflict observed in whole genome data
Richard H. Adams, Todd A. Castoe, and Michael DeGiorgio, “PhyloWGA: chromosome-aware phylogenetic interrogation of whole genome alignments,” Bioinformatics, 37(13), 1923–1925 (2021).
phylogenomics, is adding another type of controversy never seen before in molecular phylogenetics: highly supported contradictory results.
Gonzalo Giribet, “Genomics and the animal tree of life: conflicts and future prospects,” Zoologica Scripta, 45: s1, pp 14-21 (2016)
Phylogenomic conflict, where gene trees disagree about species tree resolution, is common across genomes and throughout the Tree of Life.
Caroline Parins-Fukuchia, Gregory W. Stull, and Stephen A. Smith, “Phylogenomic conflict coincides with rapid morphological innovation,” Proceedings of the National Academy of Sciences (PNAS), 118, No. 19: e2023058118 (2021).
Elizabeth Pennisi "Shaking up the Tree of Life" Science 18 Nov 2016: Vol. 354, Issue 6314, pp. 817-821
E. V. Koonin "The Logic of Chance: The Nature and Origin of Biological Evolution" (2012) : 8
نماذج أخرى لهذه التضاربات تم ذكرها فى كتاب تصميم الحياة: التصنيف و التفسير t.ly/v31D
بل و يقر التطوريون أن زيادة البيانات و المعلومات لا يحل المشكلة بل بالعكس يزيد من التضاربات و أنها مشكلة شائعة و بالتبعية لا يمكن للتطور هنا أن يهرب بزعم أننا عندما نعرف أكثر ستحل المشكلة لأن الواقع هو أنه كلما نعرف أكثر تزيد المشكلة و يتضح أنها قاعدة و ليست استثناء
Phylogenomics, the use of large-scale data matrices in phylogenetic analyses, has been viewed as the ultimate solution to the problem of resolving difficult nodes in the tree of life. However, it has become clear that analyses of these large genomic data sets can also result in conflict in estimates of phylogeny...Phylogenomics seemed poised to fulfill this promise to resolve the tree of life. However, analyses of large data matrices have sometimes yielded incongruent topologies, emphasizing that data collection alone is not sufficient to reach this goal.
Reddy et al., “Why Do Phylogenomic Data Sets Yield Conflicting Trees? Data Type Influences the Avian Tree of Life more than Taxon Sampling,” Systematic Biology, 66(4): 857-879 (2017).
"Incongruence between phylogenies derived from morphological versus molecular analyses and between trees based on different subsets of molecular sequences has become pervasive as datasets have expanded rapidly in both characters and species...phylogenetic conflict is common and frequently the norm rather than the exception"
"Understanding phylogenetic incongruence: lessons from phyllostomid bats," Biological Reviews of the Cambridge Philosophical Society (2012), 87:991–1024.
"A major challenge for incorporating such large amounts of data into inference of species trees is that conflicting genealogical histories often exist in different genes throughout the genome. Recent advances in genealogical modeling suggest that resolving close species relationships is not quite as simple as applying more data to the problem...evolutionary trees from different genes often have conflicting branching patterns"
James H.Degnan and Noah A.Rosenberg "Gene tree discordance, phylogenetic inference and the multispecies coalescent" Trends in Ecology & Evolution Volume 24, Issue 6, June 2009, Pages 332-340
"the more we learn about genomes the less tree-like we find their eolutionary history"
Bapteste et al., 2013, Networks: expanding evolutionary thinking, Trends in Genetics, 29, pp 439-441
Making the Impossible Possible: Rooting the Tree of Placental Mammals," Molecular Biology and Evolution (2013).
Hervé Philippe et al., "Resolving Difficult Phylogenetic Questions: Why More Sequences Are Not Enough" PLoS Biol. 2011 Mar; 9(3): e1000602.
Antonis Rokas & Sean B. Carroll, “Bushes in the Tree of Life,” PLoS Biology, Vol 4(11): 1899-1904 (Nov., 2006)
و الورقة الأخيرة تحديدا أشارت الى أن احدى عمليات بناء شجرة الحياة اضطرت الى حذف 35% من الجينات لأنها أعطت نتائج لا تتفق مع العلاقات التطورية المفترضة و مراجعات أخرى أشارت الى أن أنصار التفكير الشجرى يتجاهلون المشاكل على أنها مجرد ضوضاء (عدم دقة) فى البيانات مع أنها قد تكون اشارت "تطورية" هامة
A tree-thinker may choose to ignore conflicting signal as if it was noise, even if legitimate evolutionary events underlie it
Bapteste, E., Susko et al., (2005). Do orthologous gene phylogenies really support tree - thinking?. BMC Evolutionary Biology, 5(1), 33.
لدرجة أن احدى الأوراق أوصت فعليا باستخدام جزء من البيانات لا كلها لتجنب تضارب الأشجار الجينية و التركيز على مجموعة الجينات التى تتواءم مع العلاقات التطورية phylogenetically informative
Our results indicate that the subset of genes with strong phylogenetic signal is more informative than the full set of genes, suggesting that phylogenomic analyses using conditional combination approaches, rather than approaches based on total evidence, may be more powerful
Leonidas Salichos & Antonis Rokas "Inferring ancient divergences requires genes with strong phylogenetic signals" Nature volume 497, pages327–331 (2013)
لاحظ أن الورقة السابقة كانت محاولة لايجاد شجرة تطورية لعلاقات أنواع فطريات الخميرة مع بعضها البعض فأعطى كل جين سيناريو تطورى مختلف...نحن لا نتحدث عن علاقات عائلات مختلفة مع بعضها البعض بل فقط عن علاقات الخميرة...تذكر هذا عندما يحدثك التطورى بمنتهى الثقة قائلا "أوه ماى داروين...لدينا ترتيب شجرى ممتاز للكائنات"...لديهم ترتيب شجرى ممتاز بعد "حذف" البيانات التى تتعارض معه
البروتينات لا تعطيك النتيجة التى تريدها ؟ لا يهم سنقول أنها غير معبرة عن التطور لأنها متعارضة مع السيناريو المفترض ثم نبحث عن شئ اخر يعطي نتيجة قريبة مما نريد...و لكن هذا لا يفسر لماذا تتعارض الروتينات مع نتائجنا...لا يهم سنحكى قصة ما: نقص فى البيانات أو تطور سريع سنجد تبريرا فى النهاية
When the researchers tried to build the new avian family tree, “we were shocked to find we couldn’t get a solid answer,” Jarvis recalls. As the consortium developed more sophisticated bioinformatics tools to analyze the genome data, they discovered that protein-coding genes by themselves were not the most reliable for building good trees. The non-coding regions within or between genes, called introns, gave better answers.
ELIZABETH PENNISI "Bird genomes give new perches to old friends" SCIENCE 12 Dec 2014 Vol 346, Issue 6215 pp. 1275-1276
We've figured out that protein-coding genes tell the wrong story for inferring the species tree. You need non-coding sequences, including the intergenic regions.
"'Big Bang' of bird evolution mapped: Genes reveal deep histories of bird origins, feathers, flight and song" Science Daily December 11, 2014
و يقر عدد من متزايد من الأوراق و الكتب بأن ممارسة استثناء البيانات المتعارضة مع التطور من الأشجار باعتبارها بيانات خاطئة من حيث المبدأ منتشرة و يحاولون تبريرها بأنها ممارسة صحيحة و منافعها أكبر من أضرارها و الحقيقة أن نفعها الوحيد هو اثبات التطور أما أضرارها فهى ببساطة تجاهل البيانات سيقود لا محالة الى نتائج خاطئة و التى هى-و ياللصدفة -صحة التطور
Caroline Parins-Fukuchi et al., "Phylogenomic conflict coincides with rapid morphological innovation" PNAS May 11, 2021 118 (19)
Scott V.Edwards et al., "Implementing and testing the multispecies coalescent model: A valuable paradigm for phylogenomics"
"Eliminate Unreliable Data" in Barbara K. Mable, Craig Moritz, David M. Hillis "Molecular Systematics"
كيف تتجاوز نظرية التطور هذه المشاكل؟
بمنتهى السهولة. هل تذكرون تعريف "الاستدلال الانتقائى" من الفصل الأول؟ سنحتاجه الان. يتم استبعاد البيانات المتعارضة مع تطور السلالات بناءا على افتراضات الداروينية فيتم اعتبار الجين ذو القصة المتوافقة مع التطور جين هام معلوماتيا phylogenetically informative أما الجين الذى يتعارض مع الفرضيات التطورية...اسف لا يوجد فى قاموس النظرية ما يتعارض معها بل اسمه جين ليس ذو قيمة معلوماتية و لا يخبرنا شيئا!!! و لعل من الأمثلة على ذلك استدلال التطوريين بأن الجين المسئول عن انزيم سيتوكروم سى متوافق مع الشجرة التطورية و لكنهم لا يذكرون أبدا أن أخيه سيتوكروم بى متعارض معها
ان الحقيقة التى لا يعرفها الكثير من أنصار التطور أن صورة الأشجار التى يستشهدون بها-الى جانب كونها لا تثبت شيئا-فان لها نسبة توافق مع البيانات تسمى Consistency Index CI معيار الاتساق. يمثل معيار الاتساق مدى امكان التوفيق بين البيانات و بين السلف المشترك مقابل الحاجة الى فرضيات اضافية من تطور متقارب و قفز جينى و تطور صفات ثم خساراتها...الخ و فى كثير من الأحيان يمكن أن تكون الشجرة الجميلة nested hierarchy أمامك ذات معيار اتساق منخفض و نسبة كبيرة من البيانات فى الواقع لا تتوافق معها مثلا43.5%
Summary of three most parsimonious trees (MPTs)... CI = 0.565
David A. Legg et al., "Cambrian bivalved arthropod reveals origin of arthrodization" Proceedings of the Royal Society B 279 (2012): 4699-4704
أو 54 أو 65%
Strict consensus of 4 equally parsimonious trees recovered from a parsimony analysis of 52 morphological characters, all characters are unordered. Tree length=214 steps, consistency index=0.35
Rodolphe Tabuce et al., "Early Tertiary mammals from North Africa reinforce the molecular Afrotheria clade" Proceedings of the Royal Society B 274 (2007): 1159-1156
ensemble consistency index = 0.36
F. Keith Barker et al., "Phylogeny and diversification of the largest avian radiation" PNAS July 27, 2004 101 (30) 11040-11045
الرئيسيات (نحن و عائلة القرود الكريمة) كائنات ذات درجة عالية من التشابه و لذا من المفترض ألا تواجه مشاكل و مع ذلك معدل الاتساق 50% فقط
consistency index = 0.548
Jonathan I. Bloch "New Paleocene skeletons and the relationship of plesiadapiforms to crown-clade primates" PNAS 2007 Jan 23; 104(4): 1159–1164.
و الحال لا يختلف كثيرا لأشجار بعض فصائل الزواحف
consistency index (CI), 0.5346
ADAM HUTTENLOCKER "An investigation into the cladistic relationships and monophyly of therocephalian therapsids (Amniota: Synapsida)" Zoological Journal of the Linnean Society, Volume 157, Issue 4, December 2009, Pages 865–891
و من الممكن تصل نسبة عدم التوافق الى أكثر من 90%...نعم من الممكن أن ترى أشجارا تصنيفية أنيقة و علاقات تطورية تتضارب معها البيانات بنسبة 90% كما فى حالة سايتوكروم b
The evolutionary history inferred using the Maximum Parsimony method [S120] for COI [Cytochrome Oxidase Subunit I]...The consistency index is 0.051612...The evolutionary history inferred using the Maximum Parsimony method [S120] for cyt b [Cytochrome b]...The consistency index is 0.065208
Shanan S. Tobe et al., "Reconstructing Mammalian Phylogenies: A Detailed Comparison of the Cytochrome b and Cytochrome Oxidase Subunit I Mitochondrial Genes" PLOS ONE (November 2010)
و قد عقبت احدى المراجعات على ذلك بقولها أن الوسائل التى يتم استخدامها لبناء هذه الأشجار مصممة بحيث تبنى لك شجرة بغض النظر عن كون هذا أفضل تمثيل للبيانات أو لا
Extracting phylogenetic signal from genomic data can be difficult, and much of the evidence for ancient relationships is inconclusive at best. Phylogenetic methods designed to build a tree will do so whether it is the best fit with the underlying data or not.
Cédric Blais and John M. Archibald, “The past, present and future of the tree of life,” Current Biology, 31: R311-R329 (2021).
بل و أشارت نفس الدراسة أعلاه الى أن السببب الحقيقى للاستمرار فى استخدام الأشجار تاريخى و ثقافى الى جانب سهولتها
The continued use of the tree of life for classification is thus as much a reflection of its practical convenience and historical and cultural inertia as it is a commitment to natural classification.
و تنصح هذه المراجعة (و اخرون) بطريقة عجيبة جدا للابقاء على شجرة الحياة و هى حساب "متوسط" الشجرة أو كما تسمى أحيانا الشجرة الاحصائية statistical/consensus tree و هى طريقة تخرجك تماما من دائرة التطبيق العملى الى الكلام النظرى فتخيل مثلا شخص وجدوا ذراعه فى القطب الشمالى و قدمه فى القطب الجنوبى فأصبح "متوسط" مكان وجوده احصائيا على الورق عند خط الاستواء فى المنتصف. هذا كلام سليم "احصائيا و حسابيا" لكنه مستحيل عمليا فلا يمكن أن يوجد متوسط احصائى للكائن فى الطبيعة خارج الحسابات النظرية على الورق و لكن يبدو أنه البعد التاريخى و الثقافى الذى يتحدث و ليس العلم.
طريقة أخرى لتجاوز المشكلة القاء الفرضيات يمينا و يسارا فاذا كان التطور يفترض أن السلالات انفصلت عن سلف مشترك قريب و بالتالى يجب أن تكون اختلافاتها قليلة يتم استبعاد الاختلافات الكبيرة من البيانات بافتراض لا دليل عليه أنها تعرضت لطفرات كثيرة فى زمن قصير و بالتالى هى بيانات غير معبرة عن التطور!!! و اذا كانت السلالات يفترض أنها انفصلت عن بعضها منذ زمن بعيد و بالتالى يجب أن تكون قد تطفرت كثيرا و اختلفت بشكل كبير يتم استبعاد التشابهات بزعم أنها نتاج سلسلة طويلة من طفرات كثيرة و لكن أدت الى نتائج متماثلة و بالتالى فهى أيضا بيانات غير معبرة عن التطور!!! و لنأخذ مثالا بسيطا من ورقة Baum et al., التى تناولناها كنموذج للأدلة الاحصائية المزعومة على السلف المشترك فى مقالة التنادد. بالنظر الى المصادر التى اعتمدتها الورقة فى اختيار عينة الصفات نجد أن الصفات تم انتقاءها خصيصا حتى يمكن الاستدلالل بها على التطور
We examined sequence variation in 18 homologous gene segments (including nearly 10,000 base pairs) that were selected for maximal phylogenetic informativeness in resolving the hierarchy of early mammalian divergence
William J. Murphy et al., "Molecular phylogenetics and the origins of placental mammals" Nature volume 409, pages614–618 (2001)
لكن المشكلة الأكبر تظهر عند النظر الى الجدول رقم 1 فى الورقة (ينصح بتنزيل ملف البى دى اف لأن الجدول فيه أوضح) نجد أن معيار الاتساق مع السلف المشترك فى 14 جين من 16 CI يتراوح بين 0.25 و 0.46 و واحد 0.51 و الاخر 0.65 باجمالى متوسط 0.395. ان كانت الجينات المنتقاة بعناية لاثبات العلاقات التطورية تبدى معيار اتساق بنسبة تعارض النشأة فى سلف مشترك بنسبة تتراوح بين 54% و 75% فى معظم الحالات و بمتوسط 60% بعد حساب الشذوذات المرتفعة فما بالك بالجينات التى لا تظهر العلاقات التطورية؟ ان العبرة هنا ليست بتأليف قصص لترقيع البيانات من طراز ربما حدث تطور متقارب أو انتقال أفقى للجينات أو تطور سريع ثم تقديم الشجرة على أنها تظهر العلاقات التطورية بل برؤية البيانات كما هى و هى أن السلف المشترك يمثل شذوذا فى البيانات و ليس قاعدة ما دونها الشذوذ (هذا على التسليم أصلا بأن التشابه الشجرى سببه سلف مشترك فعلا و ليس احتياجات وظيفية مشتركة بين نوعين مستقلين)
و بما أننا ذكرنا الانتقال الأفقى للجينات فهى فرصة لنوضح أنه نموذج اخر من الأساليب التطورية فى التعامل مع البيانات المتعارضة مع السلف المشترك...فان لم ترد تجاهل البيانات أو اطلاق مصطلح التطور المتقارب يمكنك أن تقول أن الجينات انتقلت من الكائنات أحادية الخلية الى الخلايا التكاثرية للكائنات متعددة الخلية أفقيا أو حتى بين متعددات الخلايا نفسها و هذه هى "الموضة" الجديدة بعد التطور المتقارب...فى هذه الورقة مثلا تم العثور على جينات تشفر لمجموعة انزيمات موجودة فى الفقاريات و بعض أنواع البكتيريا و غائبة من كل ما بينهما
Thomas F. M. Cummings et al., "Citrullination was introduced into animals by horizontal gene transfer from cyanobacteria" Molecular Biology and Evolution, November 2021
ما هو الحل؟ سهلة جدا...سنرسم سهما بعرض شجرة التطور من البكتيريا الى الحيوانات...مهلا هناك مشكلة...السلف المفترض الذى رسمنا سهم الانتقال اليه توجد كائنات تتفرع منه تملك الانزيمات و أخرى لا تملكها...و هل هذه مشكلة للتطور يا رجل؟ سنفترض أن بعض الكائنات التى انحدرت من السلف احتفظت بالجينات و أخرى فقدتها

الشكل رقم 5 فى الورقة أعلاه مع اضافة تظليل
هل هناك أى دليل على هذا الكلام؟ طبعا...الدليل هو أن التطور حقيقة علمية و يجب أن يتم ترقيع البيانات وفقا لها مهما كلفنا ذلك من أسهم بالطول و العرض و الزاوية و خطوط مستقيمة و أخرى منقطة ترمز للحصول على الجينات ثم فقدها أو ادعاء فرضيات لا دليل عليها كالتطور المتقارب و النشوء من الصفر و ضوضاء البيانات...الخ...لقد أصبح الأمر شبيها بنموذج مركزية الأرض فى القرون الوسطى عندما كانوا يواصلون رسم المزيد من الدوائر حول الأرض لاستيعاب المدارات المختلفة حتى لا يعترفوا بأن الأرض ليست هى المركز!!! لا تقل هذا بالتأكيد لديهم أسلوب علمى للتأكد من هذه الحالات
Sequence analysis identifies horizontal transfers when a novel protein abruptly appears in a phylogeny with clear ancestry from the same type of protein in a taxonomically distant organism. For example, herbivory emerged rapidly among both nematode worms and beetles through the acquisition of enzymes from diverse bacteria and fungi that are capable of digesting complex plant polymers.
James A. Shapiro "Evolution Is Neither Random Accidents nor Divine Intervention: Biological Action Changes Genomes" National Association of Scholars, Academic Questions Volume 37 Issue 1 (The State of Evolution) Spring 2024
In the phylogenetic approach, each instance of topological discordance between a gene tree and a trusted reference tree is taken as a prima facie instance of LGT.
Mark A. Ragan and Robert G. Beiko, “Lateral genetic transfer: open issues,” Philosophical Transactions of the Royal Society B, Vol. 364:2241-2251 (2009).
نعم!!!هو عينه ما قرأت...عندما نجد البيانات الجينية لا تتفق مع السلف المشترك نعتبر هذا انتقالا أفقيا للجينات...هكذا عندما تتعارض البيانات مع النظرية نتهم البيانات و نبحث عن أى تأويل أو التفاف عليها و الدليل على هذا التأويل و الالتفاف هو أن البيانات تعارض النظرية!!! كم هذا رائع!!! تذكر أن التحدى التطورى قديما كان أن على من يريد نفى التطور أن يأتى بما لا يتفق مع التصنيف الشجرى فلما ظهرت البيانات التى لا تتفق مع الشجرة تم تجاهل الذى طرحوه و بدأوا فى اختراع التبريرات
We first suspected that there is horizontal transfer of mitochondrial genes by finding three striking distributional anomalies in a survey of mitochondrial gene content in angiosperms...These results establish for the first time that conventional genes are subject to evolutionarily frequent HGT during plant evolution and provide the first unambiguous evidence that plants can donate DNA horizontally to other plants
Ulfar Bergthorsson et al., "Widespread horizontal transfer of mitochondrial genes in flowering plants,” Nature, Vol. 424:197-201 (July 10, 2003).
لقد وجدنا جينات موزعة بشكل لا يتفق مع السلف المشترك و هذا دليل دامغ على أن النباتات تتبادل الجينات فيما بينها!!! الدليل على هذه الفرضية هو أن البيانات تعارض التطور!!! بامكانك النظر الى شكل رقم 4 فى الورقة السابقة لترى صورة بديعة مليئة بالأسهم العابرة للسلالات و الدليل على هذه الأسهم؟ أنها الطريقة المطلوبة لتوفيق البيانات مع التطور!!! لاحظ عنوان الورقة: اكتشفنا أن الانتقال الأفقى للجينات منتشر بين النباتات و لكن كلمة "اكتشفنا" هنا فى الواقع و بعد معرفة التفاصيل يتضح أنها اكتشفنا تعارض البيانات مع فرضية السلف المشترك و وضعنا تفسير لانقاذ الفرضية ثم تظاهرنا أن هذا التفسير هو الاكتشاف و ليس تعارض البيانات مع فرضية السلف المشترك و أشجارها (ملحوظة: لا أتهم الباحثين بالكذب عمدا و لكن مع الأسف حشو عقولهم ليلا نهارا أثناء الدراسة بأن التطور حقيقة يجعلهم يقومون بتأويل البيانات تعسفيا على مقاس هذه "الحقيقة" التى تتعارض مع البيانات)
و بنفس المنطق العقيم تخبرنا دراسة أخرى أن وجود تسلسلات متشابهة فى كائناات متباعدة تطوريا و لكن تعيش فى نفس البيئة دليل على الانتقال الأفقى للجينات...ألا يحتمل أن تكون موجودة لأنها تخدم وظيفة مناسبة للبيئة؟ طبعا لا فوثن التطور يخبرنا بأن التسلسلات تأتى أولا ثم تجد لها وظيفة لاحقا (ان وجدت) اذن فهو الانتقال الأفقى للجينات لأن التطور يتطلب هذا
Thus, hyperthermophilic bacteria were found to exhibit much higher sequence similarity to the archaea that share the same habitat than to mesophilic bacteria, likely as the result of archaea-to-bacteria LST...Likewise, our analysis of 16 bacterial genomes found a significant proportion of genes without homologs in closely related species but with homologs in distantly related taxa. These genes were likely acquired through lateral transfer.
Vicky Merhej and Didier Raoult "Rhizome of life, catastrophes, sequence exchanges, gene creations, and giant viruses: how microbial genomics challenges Darwin" REVIEW article Front. Cell. Infect. Microbiol., 28 August 2012 Sec. Molecular Bacterial Pathogenesis
لكن الملفت أكثر فى الورقة السابقة هو محاولتها لوضع رسم توضيحى لجينات الميتوكوندريا المتبادلة وفق زعمهم ليس فى البكتيريا فحسب و لكن أيضا فى بعض متعددات الخلايا كالبشر

ان وجود قدرة لدى البكتيريا على تبادل الجينات شيء و الافراط فى وضع السيناروهات كلما وجدنا تشابها لا يتفق مع شجرة التطور شيئ اخر تماما (انظر شكل رقم 6 فى الورقة السابقة) لكن أن يتم نقل الأمر الى متعددات الخلايا و بهذه الكثافة و فى جينات مفصلية للحياة كالمايتوكوندريا المطلوبة لتوليد الطاقة فان هذا هروب من البيانات أكثر منه تفسير للبيانات. و بعيدا عن الافراط فى استخدام هذا السيناريو بدون دليل على حدوثه لترقيع البيانات على مقاس التطور فان هناك طعن أصلا فى "امكان" حدوثه بهذا الشكل ابتداءا خاصة و أنه كثيرا ما يتم افتراضه لمتعددات خلايا لا يمكن أن تتزاوج أصلا و يتم وضع سيناريوهات بالغة اللاحتمالية له
William F Martin "Too Much Eukaryote LGT" Bioessays (2017);39(12)
حتى فورد دوليتل التطورى الشهير و الذى يعد أصلا من أنصار شيوع الانتقال الأفقى للجينات ينتقد الطريقة التى يتم استخدامها موضحا أن ما يحدث هو مقارنة البيانات مع شجرة معيارية تطورية مشكوك فيها أصلا ثم اعتبار ما يخرج عليها انتقال أفقى للجينات
Many studies use a false null. Attempts to measure LGT as incongruence among gene family datasets across the prokaryotes often founder on the patchy distributions and weak phylogenetic signals that characterize most of them. It is thus common to look among such datasets for strongly supported incongruence with respect to reference trees (scored as LGT), while assuming that genes that statistically ‘do not reject’ the reference topology exhibit VD.
Doolittle, W.F. 2009. The practice of classification and the theory of evolution, and what the demise of Charles Darwin’s tree of life hypothesis means for both of them. Phil. Trans. R. Soc. B 364: 2221–2228
المدهش أن دوليتل فى المقال ينتقد الخلقويين على تصويرهم للتطور كعقيدة دوغمائية بينما ما يفعله هو و زملاؤه يثبت هذا الأمر فهم يحاربون ليثبتوا شجرة الحياة رغما عن أنف الأدلة و هو يخترع قصة لا أساس لها عن سلف مشترك عبارة عن جماعة من الخلايا تمتلك جينات متنوعة تتبادلها فيما بينها و يتحدث عن المكاسب الاجتماعية و السياسية فى الحرب الثقافية ضد الخلقوين! من أين جاءت جماعة الخلايا؟ من سلف مشترك؟ اذن لم تحل شيئا فقط قمت بدفع المشكلة لمكان اخر لاعطاء نفسك مبرر لغياب الدليل. بدون سلف مشترك؟ أى أن الخلايا الحية نشأت من المادة الغير حية عدد كبير من المرات لتصنع جماعة حية و هذا مستحيل
حاول أيضا أن تتذكر أن التطوريون كثيرا ما يتحججون (و فى رواية أخرى يتبجحون) بأن تفسيراتهم أفضل من القول بالتصميم لأنها the most parsimonous الأكثر اقتصادا فى الفرضيات الاضافية وفقا لنصل أوكام. واضح!!! ما لا يقوله لك التطورى و هو يتظاهر بأن هدفه فقط هو التفسير العلمى الأفضل و الأكثر اقتصادا فى الفرضيات الاضافية هو أنه بمجرد أن يزيل فرضية التصميم يضيف بدلا منها عشرات الفرضيات البلهاء الغير منطقية و الغير محتملة من سلالات شبحية و جينات تقفز يمينا و يسارا و مسارات تطورية مختلفة تصل الى نفس النتائج و سرعات متباينة لتطور الجينات و صفات تكتسب ثم تفقد ثم تكتسب...الخ و لا يأبه لا لنصل أوكام و لا لمبدأ الاقتصاد parsimony بمجرد أن حقق هدفه الفلسفى الحقيقى: ازالة التصميم.
أضف الى ذلك أننا اذا سايرنا التطوريين فى كلامهم عن شيوع الانتقال الأفقى للجينات الى درجة كبيرة (بعضهم يقول أنه حدث مثلا فى حوالى ربع الجينات المشتركة بين الثعابين والأبقار) فلا يوجد أدنى سبب لنفترض أنه قاصر على الجينات التى تعارض سيناريو التطور. لماذا لا تكون الجينات التى يحتج بها التطوريون كونها موافقة لأشجارهم المزعومة هى التى انتقلت أفقيا؟ هنا سيقول لك التطورى أن غالبية البيانات تدعم التطور لذا هو يفترض الانتقال الأفقى فقط فى ما يعارضه و هذا زعم مضحك فكما استعرضنا فى هذا المقال نسبة البينات التى تدعم التطور فى العلاقات الجينية و التشريحية ليست أبدا بالمستوى الذى يزعمونه هى فقط لا تظهر لأنه يدارونها باستمرار بمصطلحات كالانتقال الأفقى و معايير الاتساق و التطور السريع و البطئ و الجينات الغير معبرة عن العلاقات الفيلوجينية و الانتواع السريع و غيرها. اذن فمن جديد طالما سنفتح هذا الباب فلا مانع أبدا من افتراض ن بعض الجينات التى تدعم العلاقات التطورية فى الواقع منتقلة أفقيا لا موروثة رأسيا. السبب الوحيد الذى يمنع هذه الفرضية هو أن التطوريون يحاولون الحفاظ على فرضياتهم المهترئة و التخلص فقط مما يعارضها لذا يوظفون فرضيات الانتقال الأفقى بشكل انتقائى. بل ان لدينا حالات عملية ثبتت فيها أن أوهام بعض التسلسلات الداعمة للعلاقات التطورية متقلة أفقيا و ليست موروثة من سلف مشترك
Further, it is striking that for many other RNA viruses for which there is evidence for cospeciation, including GBV-A/GBV-C, arenaviruses, and hantaviruses, there are also clear examples of cross-species transmission. If these viruses evolve at the same rate as other RNA viruses, then a process other than cospeciation must have produced the remaining match between the virus and host trees. Two mechanisms seem most likely. First, if phylogenetically related host species tend to live sympatrically, then viruses will tend to jump between closely related host species, which appears to be the case for the hantaviruses (2). If this process occurs at sufficient frequency, then the host and virus trees may often match, giving a false impression of cospeciation. Second, the ability to jump species boundaries may be dependent on the phylogenetic distance between hosts, so that it is easier to establish a new infection in a closely related host species than in a more distantly related one (5, 8).
Edward Holmes “Molecular Clocks and the Puzzle of RNA Virus Origins” Journal of Virology, Vol. 77, No. 7. April 2003: p. 3893 – 3897
و اخيرا و ليس اخرا فاننا حتى اذا سلمنا جدلا بشيوع ظاهرة الانتقال الأفقى للجينات و أنها حل المعضلة فان هذا يتركنا مع السؤال البديهى: من أين جاءت كل هذه الجينات التى تنتقل يمينا و يسارا؟ ان القول بأن جينا ما كان موجودا فى كائن و انتقل الى اخر أو أن السلف المشترك لم يكن نوعا واحدا من الخلايا بل كان جماعة من خلايا متعددة الأنواع تتبادل الجينات فيما بينها و غيرها من فرضيات الانتقال الأفقى للجينات لا يعفى التطوريين من تفسير أصل كل جين من هذه الجينات التى يتم تبادلها و كل خلية من خلايا جماعة الخلايا السلف الأسطورية المزعومة و لكنه فى المقابل يسقط حقهم تماما فى تكرار دليل الأشجار المزعوم.
بل انه حتى فى حالات الكائنات وحيدة الخلية سواءا من البكتيريا (بدائيات النوى) أو الخمائر (حقيقيات النوى) و التى فعلا تكون سيناريوهات الانتقال الأفقى للجينات بينها معقولة الى حد ما لا نسلم للتطوريين أصلا بعشوائيتها اذ وجدت الدراسات أن الجينات تنتقل على منصات تضم معها اليات لدمجها فى الجينوم المتلقى و توجد انزيمات متخصصة لادخالها فى الكائن الجديد فى مواقع متخصصة مجهزة لذلك أى أن العملية أبعد ما تكون عن العشوائية و الصدفة
tyrosine recombinase as essential for their mobilization into genomic sites with a specific target site consensus sequence...Integrative and Conjugative Elements (ICEs) play a well-established role in disseminating the genetic information underlying adaptive traits. ICEs are mobile DNA (~20 Kbp to >500 Kbp in size) that contain the genes required for genomic integration, excision, and transfer via conjugation.
Andrew S. Urquhart et al., "Starships are active eukaryotic transposable elements mobilized by a new family of tyrosine recombinases" PNAS Vol. 120 | No. 15 (April 6, 2023)
تخبرنا الأبحاث التطورية ان ما يسمى الانتقال الأفقى للجينات ظاهرة شائعة فى أشجار الحياة التطورية و قد وثقت احدى الدراسات عشرات الأمثلة من جينات مشتركة بين نباتات و حشرات و فطريات و طحالب و فقاريات
Crsip A et al., "Expression of multiple horizontally acquired genes is a hallmark of both vertebrate and invertebrate genomes" Genome Biology (2015): 16:50
و لكى نترجم استنتاجات الورقة من لغة نظرية التطور الى لغة البشر العقلاء فما يقولوه هو أننا وجدنا جينات متماثلة بين كائنات لا علاقة لها ببعضها البعض فهل نعترف بأن هذا دليل يطعن فى صحة السلف المشترك؟ لا طبعا بل نقم بتأليف قصص و افتراض سيناريوهات بعيدة الحدوث جدا كحدوث عدوى لكائن و التقاط الفايروس لجين من هذا الكائن و احتفاظه به ثم قيامه بعدوى وصلت الى الخلايا التناسلية فى كائن اخر فأدخلت فيها الجين و من ثم ولد نسل جديد بجين جديد و طبعا لأن التطور رائع جدا فقد قام بدمج الجين الجديد المفاجئ فى اليات عمل الكائن الجديد...و قد تكررت هذه الصدف أقصد هذا السيناريو العلمى عشرات المرات...حتى فى الحالات التى يكون مصدر الجين فيها بكتيريا فيكون السيناريو أكثر قابلية للحدوث نجد الجين "المنتقل أفقيا" قد تم تعديله بوضع انترونات (الية تنظيمية) غير موجودة فى نسخته الأصلية فى البكتيريا...طبعا لأن التطور رائع جدا و قام بتعديله ليلائم الكائن الجديد!!!ما الدليل على حدوث كل هذا؟ الدليل هو أن التطور حقيقة طبعا فعندما نجد جينات متعارضة معه نقم بتأليف سيناريوهات و قصص عن كيفية وصول هذه الجينات الى هذه الكائنات أو تطورها عدة مرات بشكل مستقل بلا سلف مشترك مثلا:
لقد درسنا جينوم الطيور و أثبتنا السلف المشترك و صنعنا شجرة تطورية ممتازة متناسقة...فقط بعد أن اضطررنا لافتراض أن الكثير من السمات نشأت بدون سلف مشترك أكثر من مرة أو نشأت من سلف مشترك و لكن الدليل مفقود بسبب عدم اكتمال البيانات
representing independent lineages of diverse and convergently evolved land and water bird species...confirm independent gains of vocal learning...Even with whole genomes, some of the earliest branches in Neoaves proved challenging to resolve, which was best explained by massive protein-coding sequence convergence and high levels of incomplete lineage sorting that occurred during a rapid radiation after the Cretaceous-Paleogene mass extinction event about 66 million years ago.
ERICH D. JARVIS "Whole-genome analyses resolve early branches in the tree of life of modern birds" SCIENCE 12 Dec 2014 Vol 346, Issue 6215 pp. 1320-1331
there is increasing evidence that loci can have conflicting evolutionary histories (so that their phylogenetic trees are topologically different) because of many biological causes, including incomplete lineage sorting (ILS), a process that is especially common in rapid radiations,
SIAVASH MIRARAB et al., "Statistical binning enables an accurate coalescent-based estimation of the avian tree" SCIENCE 12 Dec 2014 Vol 346, Issue 6215
ان البحث عن مسائل مثل التطور المتقارب الجزيئى molecular convergence أو حتى التطور المتقارب فحسب و مثل الانتقال الأفقى للجيناتHorizontal/Lateral Gene transfer و تقارب أفرع أشجار الحياة long branch attraction و غيرها من الفرضيات الموضوعة لمحاولة تفسير لماذا الأشجار التطورية لا تتطابق مثل incomplete lineage sorting حيث تعطى الجينات نتائج متضاربة أو تشير الى تاريخ انفصال عن السلف المشترك لا يتطابق مع ظهور السلالات فى الأحافير فيفترضون أن السبب نقص البيانات أو عملية تطور سريعة لم تترك أثرا و introgression حيث يفترض مرور جينات من الخلف الى السلف فى عكس الاتجاه الطبيعى بسبب تزاوج بين سلالات مختلفة و غيرها من الفرضيات يوضح لك كم التضارب و انعدام الاتساق فى الأشجار بعكس الزعم التطورى اذ أن هذا التضارب هو سبب ابتكار هذه الفرضيات ابتداءا بل ان هناك برامج كمبيوتر مصممة لفرض الافتراضات التطورية على البيانات و توفيقها قسرا مع شكل الشجرة و مع ذلك يخرج علينا التطوريون مؤكدون اتساق الأشجار و لعله من المثير للسخرية أن هذا حرفيا هو تنبؤ رتشارد دوكنز بصحة التصميم فى كتابه "أعظم استعباط على وجه الأرض: أدلة التطور" اذ بعد أن انتهى من فاصل الكذب الذى أكد فيه اتساق الأشجار قال بالحرف أن اتساق الأشجار هو أكبر دليل على حقيقة التطور بينما عدم اتساقها قد يشير الى مصمم يقوم بوضع الجينات الأفضل فى المكان الأفضل بغض النظر عن التسلسل التطورى
"What turns this into extremely powerful evidence for evolution is that you can construct a tree of genetic resemblances separately for each gene in turn. And the important result is that every gene delivers approximately the same tree of life. Once again, this is exactly what you would expect if you were dealing with a true family tree. It is not what you expect if a designer had surveyed the whole of the animal kingdom and picked and chosen - or 'borrowed' - the best proteins for the job, wherever in the animal kingdom they might be found."
Dawkins "The Greatest Show on Earth: The Evidence for Evolution" (2009) p. 321-322
لاحظ أننا ناقشنا سابقا أن اتساق الأشجار ليس دليلا على التطور لأنها أشجار تصنيفية و التصنيف لا يدل على أن التصنيفات تتحول الى بعضها البعض و لكن ما بالك بأن التصنيف نفسه فشل تطوريا و و تضارب و لا يمكن لدوكنز و باقى كتائب الترويج للتطور أن يكونوا يجهلون هذا الكلام اذ كتاب دوكنز الذى يحتوى على الاقتباس السابق صادر سنة 2009 و حتى بعد ذلك بكثير لا تزال جوقة البروباجندا التطورية تكرر نفس الاستدلال برغم أن مشاكله بدأت تظهر من منتصف التسعينات عندما ظهرت نتائج تحليل سايتوكروم بى الغير متسقة مع شجرة التطور و بعض مشاكل سايتوكروم سى كذلك و ظهرت معها الترقيعات الشهيرة (هذا يتطور بسرعة و ذاك ببطء/انتقال أفقى/انتواع سريع/المجموعة المتطورة لم تحتوى على كل الجينات...الخ) و منذ ذلك الحين واصلت التضاربات التراكم مع ازدياد عدد الجينات المدروسة
The theoretical problem of assessing congruence between trees generated from different data sets is still unsolved. However, in practice, we find that incongruence between molecular trees (generated from different data sets or by different analytical methods) is as striking or pervasive as is incongruence between trees generated by morphologists in the long history of their discipline...As morphologists with high hopes of molecular systematics, we end this survey with our hopes dampened. Congruence between molecular phylogenies is as elusive as it is in morphology and as it is between molecules and morphology....Partly because of morphology's long history , congruence between morphological phylogenies is the exception rather than the rule. With molecular phylogenies, all generated within the last couple of decades, the situation is little better. Many cases of incongruence between molecular phylogenies are documented above; and when a consensus of all trees within I % of the shortest in a parsimony analysis is published , structure or resolution tends to evaporate
Colin Patterson, David M. Williams, and Christopher J. Humphries "Congruence Between Molecular and Morphological Phylogenies" Annual Review of Ecology and Systematics (1993) , Vol. 24:153-188
the mitochondrial cytochrome b gene implied a similarly absurd phylogeny of mammals, regardless of the method of tree construction. Cats and whales fell within primates, grouping with simians (monkeys and apes) and strepsirhines (lemurs, bush-babies and lorises) to the exclusion of tarsiers. Cytochrome b is probably the most commonly sequenced gene in vertebrates, making this surprising result even more disconcerting...More intriguingly, another mitochondrial gene coding for a neighbouring component of this electron transport chain (cytochrome c oxidase subunit II) simultaneously experienced a doubling of its evolutionary rate, and phylogenetic analysis of this sequence yields a similarly bizarre tree. The most reasonable conclusion is that this is no coincidence: rather, these functionally related parts of the mitochondrial genome have undergone concerted, adaptive evolution and lost their historical signal.
Michael S.Y. Lee "Molecular phylogenies become functional" Trends in Ecology & Evolution VOLUME 14, ISSUE 5, P177-178, MAY 01, 1999
Some twenty years ago, the foundations of the tree of life were shaken by the realization that prokaryotic genomes are comprised of genes with different evolutionary histories...The subsequent explosion of whole-genome sequencing in the late 1990s and the rise of comparative genomics was expected to validate and solidify Woese’s tree. These hopes were short-lived. Phylogenetics soon showed that different genes within the same genome could yield very different tree topologies, and even closely related organisms were found to differ substantially in gene content.
Cédric Blais and John M. Archibald, “The past, present and future of the tree of life,” Current Biology, 31: R311-R329 (2021).
و مع كل تضارب جديد سيناريو جديد و فرضية جديدة فى نظرية تتباهى دائما بأنها الأكثر اقتصادا فى الفرضيات parsimony وفقا لنصل أوكام لأنها أزالت فرضية المصمم...أزالته و وضعت مكانه عشرات الفرضيات الأخرى! من 1993 و كهنة التطور يكذبون و يقولون الأشجار متسقة بل و يروجون للسيناريوهات التى يخترعونها على أنها حقائق (سايتوكروم سى و بى تطورا بسرعة معا...يا محاسن الصدف ازداد معدل الطفرات فيهما و/أو تم الوصول الى طفرات بعينها بالتوازى فى كليهما حتى يستمر عملهما المتناسق فى سلسلة نقل الالكترونات فيحفظ الانتخاب الطبيعى هذه التغيرات)
[يقول رتشارد دوكنز التطورى الشهير أن أفضل طريقة لنفى الخلق هى الأشجار التطورية التى تترتب فيها الكائنات...لكن يبدو أنه نسى أن يقول أن هذا الترتيب يختلف وفقا للبيانات المستخدمة (من جينات أو بروتينات أو صفات تشريحية) لكن الأهم من ذلك أنه حتى مع تجاهل التضارب فترتيب الكائنات بشكل هرمى أنيق لا يعنى أنها تتحول الى بعضها البعض خاصة فى ظل العجز التام لكل الاليات المقترحة كما وضحنا فى أيقونات التطور: التطور الصغروى t.ly/lfKD و داروين يتقهقر t.ly/Xq91 و أيقونات التطور: الانتخاب الطبيعى t.ly/Yhuv و أيقونات التطور: ملايين السنين t.ly/AtuF و الشبكات النمائية t.ly/dHkK و الهندسة اللاتكيفية t.ly/KKqP و أوردنا اعتراف علماء البيولوجيا التطورية أنفسهم بغياب الاليات فى مؤتمر 2016 t.ly/1H91 هل اذا تجاهلنا تضارب الأشجار مع بعضها و افترضنا تماثلها كلها فان هذا دليل حاسم على التطور؟ ببساطة لا لأن مجرد القدرة على تصنيف الأشياء بشكل تصاعدى لا يعنى بالضرورة أنها انحدرت من بعضها. اننا قادرون على تصنيف الكثير من عائلات المنتجات و البرامج بشكل شجرى تصاعدى كأنظمة التشغيل و معالجات المعلومات processors و غيرها و مع ذلك لا يعطى هذا مبررا لافتراض أن ملايين الأخطاء القابلة للحدوث فى خطوط الانتاج أو البرمجة تراكمت و تم اصطفاء بعضها من قبل المهندسين لانتاج الجيل التالى و قد نبه مايكل دنتون على هذا الأمر فى كتابه التطور نظرية ما تزال فى أزمة t.ly/xh4Z عندما أشار الى قيام بعض العلماء من غير الخلقويين بترتيب الكائنات و التصانيف بشكل مثالى مع تجاهل تام للعلاقات التطورية التى تفرض شذوذات فى الترتيب بسبب بعض العوامل كالتطور المتقارب المزعوم و هو ما استفز التطوريين الى حالة من الهستيريا بسبب معرفتهم بأهمية هذه الحجة لهم فوصفوا هذا الأمر بأنه تطرف تصنيفى Radical Cladism و اعلان لموت داروين و حاربوه بشدة و و فى عام 2018 قام أحد خبراء تكنولوجيا المعلومات باستخدام ما يسمى "مقياس الاعتماد" dependency graph على البيانات الجينية لعدد من الكائنات الحية و هو مقياس يستخدمه مهندسو البرمجيات و الالكترونيات لتحديد العناصر التى تعتمد على بعضها البعض و بالتالى يجب استخدامها معا دائما لينتج منها ترتيب شجرى للكائنات الحية لا يعتمد على العلاقة التطورية المفترضة بل على العلاقة الوظيفية المرصودة بين الجينات موضحا بذلك التدليس التطورى فى زعم احتكار التطور لفكرة الترتيب الهرمى بل و مثبتا أن الترتيب الوظيفى هو الأفضل اذ كان توافق البيانات الجينية مع نموذجه الذى أسماه الرسم البيانى للحياة بدلا من شجرة الحياة أفضل بكثير من مستوى توافقها الملئ بالتضاربات مع أشجار الحياة التطورية
All marine species depend on a marine module, and the echolocating species depend on the echolocation module. The dependency graph is essentially a tree with extra flexibility; the modules can explain genes shared between species thought to be only distantly related by common descent. A module is not restricted to reusing code from a single source, but can freely reuse from multiple sources. Compare this to common descent where each species must almost exclusively draw from a single source: its ancestral species...The predictions of the dependency graph hypothesis set out in this paper have been shown to be correct. The biological data was a better fit to a dependency graph than to a tree. The data produced by a simulated process of common descent was a better fit to a tree than to a dependency graph. The data produced by a compiler was both a better fit to a dependency graph than a tree, and a better fit to a tree than to the null model. The inferred biological dependency graphs contained were not simply the tree of life with a few additions, but instead contained many additional modules.
Winston Ewert "The Dependency Graph of Life" Bio-Complexity Vol. 2018
و لتقريب الفكرة فان الكثير من برامج الحاسب الالى لتعمل تحتاج الى تنزيل/تثبيت مكتبات من المكونات/البرمجيات الأخرى فمثلا قد تحاول تشغيل برنامج فيطالبك بتثبيت مكتبات visual c على سبيل المثال أو يقوم بتثبيتها كجزء من عملية تثبيت نفسه لأنه يعتمد عليها فى العمل و ليس لأنه "انحدر/تطور" منها أو من سلف يمتلكها...بنفس الفكرة فان الرئيسيات مثلا تمتلك مجموعة من الصفات و الجينات ليس لأنها انحدرت من سلف مشترك و لكن لأن هذه هى حزمة المكونات المطلوبة لصناعة الرئيسيات...و تشترك الرئيسيات مع الثدييات فى حزم من المكونات ليس لأنها انحدرت من الثدييات و لكن لأن "برمجيات/قواعد بيانات" الرئيسيات يتطلب عملها مكتبات المكونات الموجودة لدى الثدييات و هكذا الطريف فى الموضوع أن حجة التصنيف على عوارها و فى ظل غياب الاليات التطورية هى أصلا حجة استنباطية عقلية و ليست علمية و نحن هنا لسنا بصدد نقد الحجج العقلية مطلقا و لكن ليرحمنا التطوريين و على رأسهم دوكنز ذاته من الاصرار على أن التطور علم و ليس فلسفة]
الجينات اليتيمة
The probability that a functional protein would appear de novo by a random association of amino acids is practically zero…. creation of entirely new nucleotide sequences could not be of any importance in the production of new information.
Jacob F. Evolution and tinkering. Science. 1977; 196: 1161-1166
ان احتمال نشأة بروتين وظيفى من الصفر باجتماع عشوائى للأحماض الأمينية صفر...انشاء تسلسلات جديدة لا يصنع معلومات
فرانسوا جاكوب - من أهم علماء التطور فى القرن العشرين
the consensus view was that virtually all genes were derived from ancestral genes , with François Jacob famously remarking in a 1977 essay that “the probability that a functional protein would appear de novo by random association of amino acids is practically zero"
Stephen Branden Van Oss,Anne-Ruxandra Carvunis "De novo gene birth" PLOS Geenetics Published: May 23, 2019
لقد كان الظن السائد أن أى جين يجب أن يأتى من سلف و أن احتمال نشأة جين جديد تماما صفر
-من التنبؤات البائسة لنظرية التطور
a glaring proportion of gene families is simply not shared between most cellular lineages
Watson, A.K. et al. 2021. “Retracing lineage history: time to emphasize genetic turnover,” Trends in Microbiology 29:P957-958.
نسبة ضخمة من عائلات الجينات غير مشتركة بين الخطوط التطورية
-من مراجعة للدراسات سنة 2021
الان مشكلة أخرى. اذا كانت تسلسلات الحمض النووى الجديدة تنتج من طفرات فى أخرى قديمة فماذا عن التسلسلات التى لا يوجد لها أى شبيه فى أى كائن اخر؟ لفترة طويلة من الزمن كان السيناريو التطورى هو أن الجينات الجديدة تنشأ من تضاعف/تكرار جينات قديمة بالخطأ أثناء النسخ ثم تبدأ الطفرات فى العمل على هذه النسخ الجديدة و قد تلقى هذا النموذج ما تلقاه من النقد سواءا بسبب احتمال الحدوث أو بسبب أضراره حال الحدوث أو بسبب أن أكثر الكائنات تطورا و تعقيدا ليست هى من تمتلك أكبر الجينومات حجما و غيرها من الأسباب
Jerry Bergman "Does gene duplication provide the engine for evolution?" Journal of Creation 20(1):99–104, April 2006
Williams, A., Copying confusion: does duplication of existing DNA help evolution? Creation 25(4):15, 2003
Behe, M.J. and Snoke, D.W., Simulating evolution by gene duplication of protein features that require multiple amino acid residues, Protein Science 13:2651–2664, 2004; p. 2652
كما جرت العادة استمر التطوريون فى تصوير هذه الاعتراضات على أنها اعتراضات خلقوية و أن الية التضاعف الجينى صحيحة بل كانوا يؤكدون أن التطور العلمى سيثبت هذا الأمر كأحد أحجار الزاوية للبيولوجيا التطورية
Jianzhi Zhang "Evolution by gene duplication: An Update" Trends in Ecology and Evolution 6 (June 2003): 297
و كعادة التنبؤات العلمية البائسة لنظرية التطور كالحمض النووى الخردة و الأعضاء الغير وظيفية و غيرها أتى العلم ليثبت عكسها تماما. تم اكتشاف هذه الجينات "اليتيمة" التى لا سلف لها ORFan Open Reading Frames و لم تنشأ بالتطور من جين اخر بتكرار النسخ أو بدونه تؤدى وظائف حيوية فى الكثير من الكائنات
Comparative genome analyses indicate that every taxonomic group so far studied contains 10-20% of genes that lack recognizable homologs in other species...Recent studies in basal metazoans such as Nematostella, Acropora and Hydra have shed light on the function of these genes, and now indicate that they are involved in important species-specific adaptive processes....We propose that taxon-specific genes drive morphological specification, enabling organisms to adapt to changing conditions...Every Eukaryotic genome contains 10-20% of genes without any significant sequence similarity to genes of other species, these are classified as “orphans” Or “Taxonomically restricted genes” TRGs...such genes have arisen in the genomes of every group of organisms studied so far
Konstantin Khalturin et al., "More than just orphans: are taxonomically-restricted genes important in evolution?" Trends in Genetics, VOLUME 25, ISSUE 9, P404-413, SEPTEMBER 01, 2009
و ضع خطا تحت وظائف حيوية هذه لأنه مع بداية ظهور هذه الجينات حاول التطوريون الهرب من تبعاتها بزعم أنها تقوم بأدوار فرعية غير هامة. لاحظ ان هذا أصلا ليس ردا فنحن لا نسأل ما أهميتها و لكن نسأل كيف ظهرت فمن الممكن أن يعتبر أحدهم جهاز تكييف الهواء مثلا أمرا ثانويا و من الكماليات غير الهامة و لكن هذا لا علاقة له بكيف نشأ. و قد توالت الأبحا ث بعد ذلك تؤكد شيوع هذه الجينات اليتيمة من جهة و ضلوعها فى وظائف هامة و محورية للكائن من جهة أخرى
Taxonomically Restricted Genes (TRGs) are genes that are present only in a particular taxonomic unit and have no traceable evolutionary history. As the TR genes have no homology to genes from other organisms, they are fundamentally important for the study of the emergence of organismal unique traits such as morphological diversity, metabolic innovation, and pathogenicity...TRGs that are present at the species and genus levels garner special interest, because they are expected to perform a role in defining exclusive ecological adaptations of organisms to particular niches
Andrzej Zielezinski et al., "TRGdb: a universal resource for exploration of taxonomically restricted genes in bacteria"
Our finding that many of the essential lineage-specific TRGs in yeast and worm play key roles in the chromosome-segregation machinery is intriguing. Chromosome segregation is one of the most basic cellular functions; all cells must be able to carry this out correctly and efficiently.
Verster, A.J. et al. 2018. Taxonomically Restricted Genes with Essential Functions Frequently Play Roles in Chromosome Segregation in Caenorhabditis elegans and Saccharomyces cerevisiae. G3: Genes Genomes Genetics Volume 7, Issue 10, 1 October 2017, Pages 3337–3347
we determined the essential Caulobacter genome at 8 bp resolution, including 1012 essential genome features: 480 ORFs, 402 regulatory sequences and 130 non-coding elements, including 90 intergenic segments of unknown function...Of the 480 essential Caulobacter ORFs, 38% are absent in most species outside the α-proteobacteria and 10% are unique to Caulobacter
Beat Christen et al., "The essential genome of a bacterium" Molecular Systems Biology Volume 7 Issue 1 1 January 2011
لاحظ فى المصدر الأخير كيف أثناء محاولتهم تحديد الحد الأدنى من الجينات لأحد أنواع البكتيريا وجدوا 90 تسلسلا لا يعلمون لها وظيفة. عادة ما يتم تصنيف تسلسلات على أنها مجهولة الوظيفة بسبب عدم تشابهها مع أى تسلسل اخر لترجيح وظيفتها و بالتالى فهى تعتبر من الأيتام كذلك الى جانب نسبة الأيتام الأخرى المذكورة - و فى قلب الحد الأدنى للحياة. نفس الأمر تكرر فى العتائق بالعثور على جينات ضرورية للحياة و جزء من الحد الأدنى للجينوم و مع ذلك غير موجودة فى أنواع الخلايا الأخرى و لا حتى فى سلف مشترك مفترض للعتائق - عشرات الجينات يجب أن تظهر دفعة واحدة للكائن من العدم حتى يعيش فى بيئته
76 (~17%) either classified as “function unknown” or “general functional prediction only.” The latter two categories are hereby collectively referred to as “poorly characterized.”...The final set of essential genes are specific or largely specific to the Sulfolobales, most of which have uncharacterized functions...The final set of essential genes are specific or largely specific to the Sulfolobales, most of which have uncharacterized functions
Changyi Zhang et al., "The essential genome of the crenarchaeal model Sulfolobus islandicus" Nature Communications volume 9, Article number: 4908 (2018) (Fig. 5)
من الأمثلة الأخرى لهذا الأمر أيضا هو تجربة كريج فنتر الشهيرة التى يزعم البعض أنها صنعت خلية و لكنها فى الحقيقة أخذت أبسط نوع من الخلايا بأصغر جينوم و حذفت منه بعض الجينات ليس الا و لكن ما علاقة هذا بالموضوع؟ علاقته أن حتى هذا الشكل البسيط البدائى من الخلايا يحتوى على عشرات الجينات التى لا يمكن حذفها لضرورتهم لحياته و مع ذلك لا شبيه لهم فى أى كائن اخر فلا هو أخذهم من سلف و لا أعطاهم لخلف
J. CRAIG VENTER et al., "Design and synthesis of a minimal bacterial genome" SCIENCE 25 Mar 2016 Vol 351, Issue 6280
عشرات الجينات فى هذا المثال فقط جاءوا من العدم ثم ذهبوا اليه و قائمة الأمثال تطول بل فى دراسة شملت 16 سلالة من البكتيريا الزرقاء cyanobacteria وجد العلماء حوالى 13800 جين يتيم موزعة بين السلالات بمعدل حوالى 860 جين يتيم لكل سلالة
We found that of the 21238 distinct CLOGs identified across all cyanobacterial strains, the majority (approximately 65%) consist of single genes that have no likely ortholog in any other considered strain.
Christian Beck et al., "The diversity of cyanobacterial metabolism: genome analysis of multiple phototrophic microorganisms" BMC Genomics volume 13, Article number: 56 (2012)
و فى دراسة أخرى شملت أكثر من 60 ألف سلالة من البكتيريا اكتشف الباحثون أن حوالى 100% منها تضم جينات خاصة غير موجودة فى الباقين و بالتالى غير موجودة فى أى سلف يمكن افتراضه
Andrzej Zielezinski et al., "TRGdb: a universal resource for exploration of taxonomically restricted genes in bacteria " See Table 1
لقد وجدت دراسات مختلفة مئات الجينات المستجدة فى الأنواع الاجتماعية لتساعدها على هذا السلوك (التعاون) الذى نشأ 11 مرة بشكل مستقل (منهم 6 مرات فى النحل) و كالعادة بدأت استراتييجية التفسير عن طريق "اطلاق الأسماء" فبعضها تمت نسبته الى التطور السريع rapid و أخرى الى المتسارع accelerated و ثالثة الى النشوء من الصفر de novo مع ملعقتين من التطور المتقارب convergent لتبرير ظهور هذه الجينات فى أنواع منعزلة تطوريا.
S. Hollis woodward et al., “Genes involved in convergent evolution of Eusociality in bees” PNAS 108, no. 18 (2011): 7472
Karen M. Kapheim et al., “Genomic Signatures of evolutionary transitions from solitary to group living” Science 348 (2015):1139-1143
و فى حالات أخرى كانت الجينات الجديدة بالالاف و ليس المئات
We also found 9,361 proteins that are unique to A. cephalotes, representing over half of its predicted proteome. These proteins likely include those specific to ants or to A. cephalotes.
Garret Suen et al., "The Genome Sequence of the Leaf-Cutter Ant Atta cephalotes Reveals Insights into Its Obligate Symbiotic Lifestyle" PLOS Genetics February 10, 2011
Doori park et al., “Uncovering the novel characteristics of Asian Honey bee, Apis Cerana, by whole genome sequencing” BMC Genomics 16, no. 1 (2015): 1-5
Pedro Ferreira et al., “Transcriptome analyses of primitively Eusocial wasps reveal novel insights into the evolution of sociality and the origin of alternative phenotypes” Genome Biology 14 (2013): k20
ان هذه الدراسات تأخذ مشكلة الجينات المستجدة الى مستوى اخر تماما فمثلا فى الورقة الثاانية تم ايجاد جينات كثيرة مستجدة ليست فقط على مستوى النحل بل على مستوى سلالة بعينها من النحل لا تشاركها فيها سلالات النحل الاجتماعى الأخرى أى أن المسألة تجاوزت وجود سلف مشترك للحشرات الاجتماعية و غير الاجتماعية و دخلت فى مستوى عدم وجود سلف واحد للنحل الاجتماعى ذاته و الورقة الأولى تأرجحت بين كون الالف الجينات الجديدة خاصة بالنمل بشكل عام ام بسلالة النمل المدروسة بشكل خاص.
حالة أخرى تمت فيها مقارنة 7 سلالات من النمل وجدت نحو 4000 جين مستجد مهم للوظائف المختلفة فى كل سلالة لنصل الى حوالى 28 ألف (28,581) جين مستجد فى النمل!!! لكن الألطف أن 64 منها فقط هى المشتركة بين السبع سلالات بينما 12,054 جين أى نحو 42.2% قاصرة على سلالة دون الباقين lineage-specific genes (موزعة ما بين هذه و تلك و ليست كلها فى واحدة) أى أننا حتى لا نستطيع افتراض سلف مشترك واحد للنمل الاجتماعى ورثت منه كل هذه الجينات ناهيك عن وجود "تطور" فى العناصر التنظيمية لألفين جين اخرين بين الحشرات الاجتماعية و غير الاجتماعية و طبعا الحل: لابد أن النمل "تطور بسرعة"
Daniel Simola et al., “Social insect genomes exhibit dramatic evolution in gene composition and regulation while preserving regulatory features linked to sociality” Genome Research 23 (2013): 1235
حتى لو افترضنا أن الجينات متشابهة و بالتبعية موروثة من سلف ما فان هذا أصلا لا يفسر نشأتها فى السلف و لكن تشعبها الشديد و اختلافها بين أنواع النمل ذاتها يطعن حتى فى وجود سلف مشترك للنمل الاجتماعى و طبعا كما جرت العادة يتم افتراض تطور بعض الجينات بسرعة و أخرى بمنتهى السرعة و تجنيد أخرى لوظائف جديدة الى اخر المسميات التى لا تصلح كتفسير ثم يخرجون علينا قائلين البيانات لا تعارض أشجار تطور السلالات
لاحظ أننا نتكلم عن كائنات شديدة التقارب فهى سلالات من نفس نوع البكتيريا أو سلالات من النمل أو النحل فما بالك بما هو أبعد؟ فى كل مكان فى البكتيريا و الفطريات و الحشرات و الفئران و البشر و الحبارات و السلمندر و فى كل نوع تم دراسة الجينوم الخاص به بشكل كامل (الجينوم هو اجمالى الحمض النووى للكائن) و هذه فقط بعض الأمثلة:
Diana Ekman and Arne Elofsson, “Identifying and quantifying orphan protein sequences in fungi,” Journal of Molecular Biology 396 (2010): 396-405. doi:10.1016/j.jmb.2009.11.053. PMID:19944701.Qi Zhou, Guojie Zhang, Yue Zhang, Shiyu Xu, Ruoping Zhao, Zubing Zhan, Xin Li, Yun Ding, Shuang Yang, and Wen Wang, "On the origin of new genes in Drosophila,” Genome Research 18 (2008): 1446- 1455. doi: 10.1101/gr.076588.108. PMID:18550802Daniel N. Murphy and Aoife McLysaght, "De novo origin of protein-coding genes in murine rodents,” PLoS One 7 (2012): e48650. doi:10.1371/journal.pone.0048650. PMID:23185269David G. Knowles and Aoife McLysaght, "Recent de novo origin of human protein-coding genes,” Genome Research 19 (2009): 1752-1759. doi:10.1101/gr.095026.109. PMID:19726446Dong-Dong Wu, David M. Irwin, and Ya-Ping Zhang, "De novo origin of human protein-coding genes,” PLoS Genetics 7 (2011): el002379. doi:10.1371/journal. pgen.1002379. PMID:22102831Jing Cai et al., "De novo origination of a new protein-coding gene in Saccharomyces cerevisiae" Genetics, Volume 179, Issue 1, 1 May 2008, Pages 487–496
Kumar, A., Gates, P., Czarkwiani, A. et al. "An orphan gene is necessary for preaxial digit formation during salamander limb development" Nature Communications 6, 8684 (2015)
Geng, J. et al. 2015. Identification of the orphan gene Prod 1 in basal and other salamander families. EvoDevo 6:9
Thümecke, S. et al. The flipflop orphan genes are required for limb bud eversion in the Tribolium embryo. Frontiers in Zoology volume 14, Article number: 48 (2017)
Rakitov, R. et al. 2018. Brochosomins and other novel proteins from brochosomes of leafhoppers (Insecta, Hemiptera, Cicadellidae). Insect Biochemistry and Molecular Biology 94 (2018) 10e17.
The analysis also turned up hundreds of other genes that are specific to the octopus and highly expressed in particular tissues.
Alison Abbott "Octopus genome holds clues to uncanny intelligence" Nature (2015)
Our analyses found hundreds of coleoid- and octopus-specific genes, many of which were expressed in tissues containing novel structures
Caroline B. Albertin et al., "The octopus genome and the evolution of cephalopod neural and morphological novelties" Nature volume 524, pages 220–224 (2015)
Because the genes of the periculin family have no orthologs in eukaryotes outside the genus Hydra, they represent taxonomically restricted genes that probably originated within the genus Hydra.
Fraune, S. et al. 2010. In an early branching metazoan, bacterial colonization of the embryo is controlled by maternal antimicrobial peptides. PNAS 107(42): 18067–18072.
و الجينات اليتيمة قد تحمل أسماء كثيرة فعندما يتم اكتشافها على مستوى البروتينات مثلا تسمى singletons أى البروتينات الوحيدة/المتفردة و التى لا يتم العثور على أى شبيه لها خارج تصنيف معين تتواجد فيه و قد وجدت احدى الدراسات التى حاولت تصنيف هذه البروتينات أن 68% من أكثر من مليار و نصف مجموعة بروتينات تمثل مجموعة يتيمة/متفردة و ستلاحظ أنهم يحاولون الالتفاف حول دلالة الأمر بذكر أن "تعداد" البروتينات الغير يتيمة أكثر (93-94%) و هذا طبيعى لأنها موجودة فى كل الكثير من التصنيفات ان لم يكن كلها (تخيل الهيموجلوبين مثلا من البديهى أن يتواجد فى الكثير من التصنيفات التى تحتاج الى جزئ نقل أكسجين) لكن المشكلة تظهر عن عد عائلات/تصنيفات البروتين و ليس عد تكرار أحاد البروتينات بين تصنيفات كائنات مختلفة
As a result of clustering ∼19 billion sequences with 30% sequence identity and 90% coverage thresholds across the tree of life, we determined ∼1.70 billion clusters with 32% of clusters yielding more than one element and 68% denoting singletons (only one unique sequence within each singleton cluster). While this majority of singletons suggests the presence of a large pool of putatively novel proteins (orphan polypeptides) within the protein universe (Supplementary fig. 11), these ∼1.16 billion unique sequences comprise only ∼6% of the full set of 19 billion sequences. The fact that 544 million clusters can capture ∼94% of all known proteins illustrates the potential of deep-clustering the protein universe to accelerate protein search across the tree of life.
Benjamin Buchfink et al., "Sensitive clustering of protein sequences at tree-of-life scale using DIAMOND DeepClust" (2023)
Yves-Henri Sanejouand "On the unknown proteins of eukaryotic proteomes" Journal of Molecular Evolution volume 91, pages492–501 (2023)
[لقاء مع فيلسوف البيولوجى بول نيلسون يفصل فيه تبعات هذه المسألة بالنسبة للتطور]
[لقد كان التطوريون يعتبرون نشأة الجينات من الصفر de novo أمرا غير ممكن لأسباب معتبرة جدا منها ضرورة وجود ما يسمى باطار القراءة المفتوح الذى يسمح بقراءة الحمض النووى فلا يمكن أن ينشأ الجين عشوائيا فى أى موضع و يجب أن يكون له promoter تسلسل مخصوص يحفز بدء نسخ الجين حتى تتم ترجمته و يجب وجود ما يسمى بالبنية الكروماتينية التى تسمح بنسخ الجين و يجب أن يكون لمنتجات الجين دور ما فى عمليات الخلية ابتداءا و الأهم من ذلك الندرة الرهيبة للتسلسلات الحمض النووى الوظيفية من بين التسلسلات المحتملة مما يتطلب البدء من تسلسل وظيفى ثم تحويله تدريجيا الى اخر وظيفى لأن التسلسل غير الوظيفى سيراكم جميع أنواع الطفرات و لن يعمل عليه الانتخاب الطبيعى لتحسينه [و ستتم مناقشة مسألة ندرة التسلسلات الوظيفية بتفصيل أكبر لاحقا].ان نشأة تسلسلات من الصفر معناها اجتماع كل هذه الصدف و هو ما كان لا يرجحه التطوريون أنفسهم برغم تعويلهم الكبير على الصدف و تم وصفه بأنه كالخيمياء السحرية فى العصور الوسطى التى تحول الرصاص الى ذهب (و طبعا كما جرت العادة يتعامل التطوريون مع التطور كشئ هلامى بلا تعريف محدد يمكن أن تنسب اليه أى شئ: التطور لا يمكنه انتاج جينات من الصفر بلا أسلاف - ماذا تقول؟ وجدنا جينات تنشأ من الصفر بلا أسلاف - التطور ينتج جينات من الصفر بلا أسلاف!!! تماما كجملة جون ماينارد سميث الشهيرة لقد كنا نظن التطور لا ينتج أشكال دائرية فلما وجدنا الصوت البكتيرى عرفنا أن التطور دوائر)
the emergence of complete, functional genes—with promoters, open reading frames (ORFs), and functional proteins—from “junk” DNA would seem highly improbable, almost like the elusive transmutation of lead into gold that was sought by medieval alchemists.
Adam Siepel "Darwinian Alchemy: Human genes from non coding dna" Genome Research 19 (2009): 1693-1694
Daniele Guerzoni and Aoife Maclysaght "De Novo Origins of human genes" PLoS Genetics, 11 (November 10,2011): e1002381, 1
Jean Armenguad, Celine Bland, Joseph Christie-Oleza and Guylaine Miotello "Microbial Proteogenomics: Gaining ground with the avalanche of genome sequences" Journal of Bacteriology Parasitology S3-001 (2011)
A remarkable one-third of the predicted genes in the human genome have no identifiable homologue in any other well-characterized non-primate species, even other mammals (Dehal et al. 200L). This poorly understood genomic feature is not unique to humans (or primates), as it has been repeatedly found in the sequenced genomes of other metazoans.
Michael Lynch, The Origins of Genome Architecture, 2007, p. 49,
و طبعا التبريرات لا تنتهى: لابد أن التطور عمل بسرعة شديدة فغيرالجينات بشدة فلم نميز أسلافها - لابد أنه أنتج جينات من الخردة...الخ و لكن حتى لو افترضنا جدلا أن هناك اليات خارقة غير معروفة لانتاج هذه الجينات فلا تزال زيادة أعدادها تمثل مشكلة حقيقية للاطار الزمنى لحدوث التطور فكل جين جديد يتيم يتم اكتشافه معناه احتياج التطور لفترة أطول و أطول و هذا لا يجدى معه مجرد أن تقول ملايين لسنين مع تجاهل الوقت المطلوب لانشاء هذه الجينات و سرعة ظهور الكائنات فى الأحافير
-مايكل دنتون: التطور ما تزال نظرية فى أزمة (سد الفجوة: الخلايا و البروتينات)
-ستيفن ماير : شك داروين
لقد حاول الدراونة كثيرا الهرب من المشاكل الجينية بالقول بأن الخلط فى أشجار الحياة يحدث نتيجة لتبادل بعض الكائنات الأولية كالبكتيريا الجينات و بغض النظر أن هذا الأمر فى حد ذاته يطعن فى امكانية الاستدلال على السلف المشترك الا أن هذه الحجة سقطت تماما بوجود الاختلافات فى كائنات كثيرة لا تتبادل الجينات و بظهور الجينات اليتيمة التى لا يملكها أحد ليتبادلها مع مالكها. ان المزاعم التى تطعن فى وجود جينات يتيمة بالأساس و تزعم أنها مجرد أخطاء فى التصنيف أو نقص فى البيانات فينفيها أن الجينات اليتيمة تزيد مع زيادة البيانات و تنقيح الأساليب لا تنقص كما زعموا أما تلك التى تزعم أنها جينات متطورة من سلف لكن تطورت بشدة حتى لم نعد نميزها فتعارضها نتائج الأبحاث كما تعارضها أيضا لامنطقية فكرة أن يقرر جين أن يتطفر بمعدل جنونى فجأة بعد انفصاله عن السلف بلا سبب أو الية لذلك (لاحظ كيف أن ردودهم تتناقض مع بعضها البعض أصلا لكن المهم ان يظهر أن التطور لديه ردود حتى و ان كانت متضاربة فمرة يقولون ليست موجودة و مرة موجودة بسبب التطفر السريع لجينات سلف و مرة التطور أنشأها من الصفر de novo).
Evolution Professor: Orphans Not a Problem for Evolution
The persistent presence of orphans and TRGs in almost every genome studied to date despite the growing number of available sequence databases demands an explanation...sequence divergence of ancestral genes explains only a minority of orphans and TRGs...we simultaneously underestimated the number of orphans and TRGs while overestimating the number that originated by divergence...we can be confident that those genes without detectable similarity really are orphans and TRGs, but in turn we also know that some will have spurious similarity hits giving the illusion that they have homologues when they do not in reality...our findings are consistent with the view that multiple evolutionary processes are responsible for the existence of orphan genes and suggest that, contrary to what has been assumed, divergence is not the predominant one
Nikolaos Vakirlis et al., "Synteny-based analyses indicate that sequence divergence is not the main source of orphan genes" eLife Feb 18, 2020
انه نفس الأسلوب الرخيص الذى استخدمته النظرية للهروب من بيانات السجل الأحفورى: طالما أن البيانات تعارض النظرية اذن العيب فى البيانات لا فى النظرية. ان هذه الأبحاث تنطلق فى نفيها للجينات اليتيمة من نظرية التطور نفسها لا من بيانات أو تجارب فالمنطق هنا هو "بما أن التطور لا يسمح بكل هذه الجينات اليتيمة اذن لابد أنها ليست موجودة و يجب أن نبحث عن تفسير اخر" أى أنها لا ترفض الجينات اليتيمة بسبب بيانات حقيقية و لكن لمجرد الحفاظ على سيناريو التطور]
طالما زعمت النظرية أن أنصار التصميم لا يجدون تفسيرا للتشابهات لأنهم يتوقعون أن يحتوى كل كائن جيناته الخاصة فتضم الأبقار جينات أبقار و الثعابين جينات ثعابين.
For creationists, this must be a nightmare, for any sensible model of creationism would predict cows to have cow molecules, goats to have goat molecules, and snakes to have snake molecules.
Hartl, D. L. 2005. Better living through evolution. Harvard Magazine (November–December), pp. 22–27
طبعا هذا كلام فارغ لأن انصار التصميم لم يعارضوا يوما اعادة استخدام المكونات و الأجزاء لكن انظر الى المفارقة. نحن اليوم نجد الجينات الخاصة بكل تصنيف بل هو المعنى الحرفى لمصطلح Taxonomically Restricted Genes TRGs و هو نفس ما زعموا أنه ضد التطور و يثبت التصميم و برغم ذلك لازالت نظرية التطور تؤكد لنا أن كل شئ يسير وفق تنبؤاتها!!!
من أين أتت هذه الجينات؟ كيف حدث هذا ال De novo origination ؟ أم أنه النسخة الجينية من توازن جولد المتقطع punctuated equilibrium عندما تطلق اسما على الظاهرة و تصفها ثم تتظاهر بأن ما فعلته من توصيف عبارة عن تفسير و تسميه "الية تطورية جديدة". طبعا مجرد اطلاق اسم فخم على الظاهرة لن يحل المشكلة. يجيب الدراونة ربما نشأت من تسلسلات غير وظيفية و لكن كلمة "ربما" لا تعنى شيئا. كم طفرة مطلوبة؟ هل كل طفرة لها فائدة حتى تحدث خطوة خطوة أم يجب أن تكون متزامنة؟ ما احتمالات حدوث ذلك؟ لا توجد طفرة سحرية تحول تسلسل خردة الى تسلسل وظيفى بل يجب أن تجتمع عدة طفرات على الأقل كحد أدنى و بما أن الوظيفة لم تبدأ بعد فان حدوث أى من هذه الطفرات لن يحفظه الانتخاب الطبيعى لمجرد أنه سيكون مفيدا فى المستقبل بل قد تحدث طفرة ثم تتطفر مجددا فتفقد و بالتالى يجب أن تتزامن هذه الطفرات و هو ما يزيد من حجم المشكلة كما أن دراسات الجينوم لم تعد تترك مجالا لأن يكون شيئا ما غير وظيفيا و قد تم التفصيل فى هذا أكثر عن الحديث عن أسطورة الحمض النووى الخردة t.ly/1uuW و الجينات الزائفة t.ly/e6ze و نتائج مشروع انكود. لا بأس اذن فلنفترض وجود جينات وسيطة proto-genes بين جينات وظيفية ما أو بين التسلسلات غير الوظيفية (ان كانت موجودة أصلا) و هذه الجينات اليتيمة تماما كما افترضنا وجود حلقات انتقالية وسيطة بين الكائنات و لكن كل هذه الوسائط و الحلقات الانتقالية سواءا الحفرية أو الجينية تظل وهما لا وجود له سوى فى ادعاءات التطوريين و فرضياتهم. بل اننى أذكر جيدا أنى شاهدت لقاءا مع باحثة أجرت عدت أبحاث عن هذه الجينات الأولية و وجدت بعض ما اعتبرته جينات أولية فى مرحلة النشأة و التطور فى بعض المناطق التى يفترض عدم وجود جينات فيها و قالت أنها بهذا أمسكت بالتطور أثناء عمله و هو ينشئ الجينات من الصفر denovo ثم ذكرت أنها فى بحث تال أرادت تطفير أو محو هذه الجينات الأولية proto genes ففوجئت بتأثير سلبى على الخلية و نموها. حسنا هل اعترفت فخامة الباحثة بأن هذه ليست جينات فى مرحلة النشأة بل جينات وظيفيىة بدليل تأثر الخلية سلبا بفقدانها؟ طبعا لا يجب أن نخترع أدلة على التطور! لقد فسرت ما حدث بأنها أمسكت بالتطور بعد أن أنشأ الجينات و ليس أثناء!!! صدق من قال "عنزة و ان طارت".
ملحوظة: تم نقل مبحث التوزيع الجغرافى الى مقال مستقل: https://illadinstormrage.wixsite.com/molakhasat/post/أيقونات-التطور-أشجار-الحياة-معضلة-التوزيع-الجغرافى
الخلاصة:
بناء شجرة الحياة يتم على 3 خطوات
1-افترض وجود سلف مشترك
2-تجاهل كل البيانات التى تتعارض مع شجرتك أو ضع لها فرضيات و نماذج بلا دليل
3-استنتج من شجرتك وجود سلف مشترك و بالتالى صحة التطور
اعتقد ثم استدل التى يتهمون كل من سواهم بها هم غارقون فيها اذ تبدأ النظرية بمعتقدين تأسيسيين لا برهان عليهما
1-كل ما نشأ فهو موجود بالتطور (و هذا المعتقد ليس مبنى على الأدلة كما يصورون للناس لأن كل أدلته فى الواقع هى تأويلات تعسفية للبيانات وفق المبدأ نفسه أى أنهم يقومون بحياكة المشاهدات و التفسيرات و التأويلات على مقاس التطور ثم يعتبرونها أدلة و بعض هذه التأويلات فى منتهى السخافة و الاستحالة)
2-لا يوجد تصميم (و هذا المعتقد أيضا ليس بسبب الأدلة فكلما وجدوا شيئا مصمما قالوا بل هو "وهم التصميم" لمجرد أن نظريتهم/عقيدتهم تتطلب ذلك و كل ما زعموه من أدلة له كالحمض النووى الخردة و سوء تصميم بعض الهياكل و الأعضاء الأثرية قد تم الرد عليه و تفنيده)
بناءا على هذين المعتقدين الباطلين يتم تأويل كل المشاهدات ثم الايحاء للناس أن هذه التأويلات حقائق علمية مع تجاهل تام لكونها أصلا تأويلات مبنية على مسلمات فلسفية و عقدية باطلة بل و أحيانا يزعمون أنها تنبؤات و كم من مرة سمعت من يقول أن نظرية التطور تنبأت بامتلاك الكائنات جينات متشابهة و كأنه ليس من البديهى و المنطقى بتطور أو بدون أن تتم الوظائف المتشابهة بجينات متشابهة و كأن هذه المعلومة البدهية من ابداعات نظرية التطور!!! (ناهيك عن خطأها أصلا كما وضحنا)
تؤكد لنا مجلة نيتشر أن هناك أدلة كثيرة تدعم السلف المشترك منها قدرتنا على تصنيف الكائنات الحية بشكل شجرى و توافق الأشجار التشريحية مع الجزيئية و التوزيع الجغرافى للكائنات الحية (الاستدلال بالأحافير و التشابهات ناقشناه فى مقالات مستقلة)
Douglas Theobald "A Formal test of the theory of universal common ancestry" Nature 464 (2010): 219 - 222
كما يؤكد لنا المركز القومى لتدريس العلوم فى الولايات المتحدة الأمريكية على توافق التوزيع الجغرافى مع نمط التطور
NCSE "Evolution on Islands"
كما تؤكد لنا مؤسسة بيولوجوس Biologos الدعوية التى تتألف من علماء يقومون بدعوة المسيحيين الى الايمان بالتطور على أن الأدلة المرصودة التى تؤكد السلف المشترك كثيرة (نعم أنت لم تخطئ القراءة: نظرية التطور لديها مؤسسات دعوية كأى دين)
"The observational evidence explained by common ancestry is overwhelming"
Biologos- Common Questions: What is evolution?
و هى جهات يفترض أنها علمية محايدة غير خاضعة للأهواء و الميول و الاراء الشخصية...يبدو أنهم يقصدون أنها أدلة على التطور بعد أن حذفنا منها كل ما يتعارض معه على طريقة البروفيسور كارل جايبرسون!!!



Comments